")
")
| Issue |
OCL
Volume 33, 2026
Palm and palm oil / Palmier et huile de palme
|
|
|---|---|---|
| Article Number | 20 | |
| Number of page(s) | 16 | |
| DOI | https://doi.org/10.1051/ocl/2026011 | |
| Published online | 13 May 2026 | |
Research Article
Fruit mesocarp of the Brazil-native Licuri palm (Syagrus coronata) as a dual and sustainable source of oil and lipase for biorefinery applications☆
Nouvelle source d’huile et d’activité lipase dans le mesocarpe du Licuri (Syagrus coronata (Mart.) Becc.), un palmier indigène du Brésil, pour des applications en bioraffinage
1
UNIT, Universidade Tiradentes, Av. Murilo Dantas, 300, Farolândia, Aracaju, SE, Brazil
2
Aix-Marseille Université, CNRS, UMR7281 BIP, 13402 Marseille cedex 09, France
3
ITP, Instituto de Tecnologia e Pesquisa, Av. Murilo Dantas, 300, Bloco ITP, Farolândia, Aracaju, SE, Brazil
4
UFBA, Federal University of Bahia, Department of Chemical Engineering, Rua Aristides Novis 2, Federation, Salvador, BA, Brazil
* Corresponding authors: This email address is being protected from spambots. You need JavaScript enabled to view it.
; This email address is being protected from spambots. You need JavaScript enabled to view it.
; This email address is being protected from spambots. You need JavaScript enabled to view it.
Received:
2
January
2026
Accepted:
30
March
2026
Abstract
Licuri (Syagrus coronata) is a palm species endemic to Brazil’s Caatinga biome whose biotechnological potential remains poorly explored. While the kernel is traditionally used for oil extraction, the mesocarp is discarded. In this study, oils from licuri kernel and mesocarp were extracted and compared. Kernel oil was rich in medium-chain saturated fatty acids, notably lauric acid (≈50 % w/w), whereas mesocarp oil contained long-chain unsaturated fatty acids (oleic and linoleic acids; 55.4 % w/w). The mesocarp oil also exhibited high levels of free fatty acids, resulting from endogenous lipase-mediated hydrolysis of triacylglycerols and galactolipids. Blanching the fruit before extraction inactivated lipase activity and improved mesocarp oil recovery, yielding up to 13 % w/w compared with 40 % w/w for the kernel. Significant lipase activity was retained in the delipidated mesocarp cake after cold hexane extraction, with activities up to 375 ± 12 and 475 ± 25 U/g on olive oil and tributyrin. Partial purification yielded a lipase active over a broad pH range and on various lipid substrates, including licuri kernel oil, egg phosphatidylcholine, and vinyl propionate. These results highlight Licuri fruit as a dual source of oil and lipase, supporting its potential use in an integrated biorefinery approach.
Résumé
Le licuri (Syagrus coronata) est une espèce de palmier endémique du biome de la région du Caatinga au Brésil, dont le potentiel biotechnologique reste encore peu exploré. Alors que l’amande est traditionnellement utilisée pour l’extraction d’huile, le mésocarpe est généralement éliminé. Dans cette étude, les huiles issues de l’amande et du mésocarpe de licuri ont été extraites et comparées. L’huile d’amande est riche en acides gras saturés à chaîne moyenne, notamment en acide laurique (≈ 50 % p/p), tandis que l’huile de mésocarpe contient des acides gras insaturés à longue chaîne, en particulier les acides oléique et linoléique (55.4 % p/p). L’huile de mésocarpe présente également des teneurs élevées en acides gras libres, résultant de l’hydrolyse des triacylglycérols et des galactolipides par une activité lipasique endogène. Le blanchiment des fruits avant l’extraction inactive cette activité lipasique et améliore le rendement en huile du mésocarpe, atteignant jusqu’à 13 % p/p, contre 40 % p/p pour l’amande. Une activité lipasique significative est conservée dans le cake de mésocarpe délipidé après extraction à l’hexane froid, avec des activités atteignant 375 ± 12 et 475 ± 25 U/g sur l’huile d’olive et le tributyrine. Une purification partielle a permis d’obtenir une lipase active sur une large gamme de pH et sur divers substrats lipidiques, incluant l’huile d’amande de licuri, la phosphatidylcholine d’œuf et le vinyl propionate. Ces résultats mettent en évidence le fruit du licuri comme une double source d’huile et de lipase, ouvrant la voie à sa valorisation dans une approche de bioraffinerie intégrée.
Key words: biorefinery / caatinga / palm tree / plant lipase / vegetable oil
Mots clés : bioraffinerie / caatinga / huile végétale / lipase végétale / Palmier
Contribution to the Topical Issue: “Palm and palm oil / Palmier et huile de palme”.
These authors contributed equally to this work.
© M.C. Lisboa et al., Published by EDP Sciences, 2026
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Highlights
The oil extracted from licuri mesocarp differs from the kernel oil.
It is rich in oleic and linoleic acids.
This oil is altered by an endogenous lipase activity degrading triacylglycerols and galactolipids. It can be preserved by blanching the fruit prior to oil extraction.
Lipase activity can be recovered from fresh delipidated mesocarp, providing a novel biocatalyst.
1 Introduction
The use of biomass in an integrated process enables the valorization of its production cycle through the development of bioprocesses that generate new products, co-products, and/or services (Victor et al., 2017). Such processes can be implemented through a biorefinery strategy, and it is particularly recommended in countries with high flora diversity, such as Brazil, where more than 33,200 different plant species represent 20 % of the world’s flora (Oliveira et al. 2012).
Brazil encompasses six major biomes, among which the Caatinga stands out as the only exclusively Brazilian biome, occupying approximately 10 % of the national territory and covering nearly 60 % of the country’s Northeast region (Fig. 1A) (Lisboa et al. 2020). Despite its ecological uniqueness and rich biodiversity, the Caatinga is located in one of the most socioeconomically vulnerable regions of Brazil. The northeast region is underdeveloped and requires sustainable regional progress. Addressing these structural issues requires coordinated governmental actions, public policies, and targeted development programs that promote economic diversification while preserving natural resources. In this context, valorization of native biodiversity and biotechnological innovation emerge as strategic pathways to stimulate local economies, generate income, and foster environmental sustainability (Victor et al., 2017).
Caatinga is known for its extreme climate, characterized by irregular precipitation patterns, with rainfall ranging from 400 to 800 mm per year. This region features shallow soils rich in minerals but poor in organic matter, which favors the growth of xerophytic vegetation resistant to prolonged drought. Due to these environmental conditions, species have had to undergo survival adaptations over the years, resulting in many endemic species with still unknown properties that may be of interest for the industrial sector (Silva et al., 2014; Victor et al., 2017).
Among those species, licuri (Syagrus coronata (Mart.) Becc.) is a palm tree whose fruit has a high oil content (49.2 % w/w) in its kernel (Fig. 1B), which is mostly saturated and contains more than 40 % lauric acid in its composition (Iha et al. 2014). Thus, licuri appears similar to palm kernel oil (49.8 % of lauric acid) in that respect (Sehgal and Sharma, 2020). This fruit has high socioeconomic importance in the Brazilian northeast, as some communities depend economically on its production chain. A good example of this is the work developed by the Production Cooperative of the Piedmont Region of Diamantina – COOPES (Capim Grosso/BA), which produces more than 20 foods derived from licuri endosperm, such as cookies, crackers, snacks, liquor, syrup, granola, candies, bread, ice cream, cooked licuri, milk, and oil (Aroucha and Aroucha, 2013).
As the main product of the licuri production chain, the kernel oil has already been used as a food supplement (Pereira et al., 2010; dos Santos Souza et al., 2021; Antoniassi et al., 2022; de Carvalho Alves et al., 2024; Sommo et al., 2024), raw material in the cosmetic industry in the manufacture of moisturizers (Leal et al., 2013), as a potential anti-inflammatory agent (de Oliveira Alves et al., 2024), for biosurfactant production (da Silva et al., 2024), as an antimicrobial agent (Cibele et al., 2016; Souza Dos Santos et al., 2019), as a raw material for the production of biofuels (De La Salles et al., 2010; Iha et al., 2014; Araújo et al., 2019; Aquino et al., 2022), as a growth control agent for Aedes aegypti, with larvicidal activity and oviposition inhibitory effect (Santos et al., 2017). It was also used in our research group’ studies on biocatalysis applied to hydrolysis reactions (Rodrigues et al., 2021) and esterification (da Silva et al., 2022).
Generally, the licuri production chain first aims for the ripe fruit harvest, followed by kernel (endosperm) separation and oil extraction. Endosperm oil extraction generates several co-products and waste (e.g., delipidated endosperm cake, the endocarp, and mesocarp; Fig 1B.) that could provide an additional value to this production chain.
Some research is already examining the potential use of these co-products and waste. The delapidated kernel cake has been used as a food supplement for lambs (Bagaldo et al., 2019) and as a substrate for submerged fermentation for the production of multienzyme extracts of industrial relevance (Ramos et al., 2025; Ramos et al., 2025). The endocarp has been used in the production of biochar, and its application as a natural fertilizer has been proposed (Santos et al., 2020). It could also be used as an adsorbent agent for removing diclofenac sodium from water (de Souza dos Santos et al., 2020). The mesocarp was utilized in the production of energy through its burning (Scalet et al., 2019), and the slow pyrolysis of both mesocarp and endocarp was employed to produce a fumigant agent against pests of stored grains (Mattos et al., 2021).
Although there is limited information about licuri in the literature, more knowledge is available about species that belong to the same family (Arecaceae) and tribe (Cocoseae). Moreover, from sequencing of the chloroplast genome by Áquila et al. (Áquila et al., 2018; Francisconi et al., 2023), it was shown that licuri (Syagrus coronata) has a phylogenetic similarity with coconut (Cocos nucifera) and oil palm (Elaeis guineenses), with genome identities of 98.9 % and 95 %, respectively. One peculiar point of interest regarding coconut and oil palm is the presence of lipolytic enzymes (Ejedegba et al., 2007; Ngando Ebongue et al., 2006) that can be isolated as crude preparations and may present potential application in biotechnology or be used for the selection of breeding lines producing oils with low acid values (Ngando Ebongue et al., 2008). Lipases (triacylglycerol hydrolases, EC 3.1.1.3) have been increasingly established as strategic biocatalysts in oleochemical industries, where they are applied in fat splitting, free fatty acid production, esterification, transesterification, interesterification, biodiesel, and biolubricant synthesis, as well as in oil upgrading processes, due to their high selectivity, mild operating conditions, and lower by-product formation. Moreover, the industrial development of these processes has been strongly supported by advances in enzyme production, immobilization, and process scale-up (Abdelmoez and Mustafa, 2014; Mustafa et al., 2023; Wancura et al., 2025; Tieves and Hollmann, 2025).
Among the lipases isolated from plant tissue, the enzyme present in the mesocarp of oil palm fruit is the best characterized, although its physiological role is still unknown. It is responsible for the intensive triglyceride hydrolysis occurring in mature oil palm fruit upon abscission and bruising, which can lead to the production of oils with unacceptable acid value (Ngando Ebongue et al., 2008). A solvent-based procedure has been established to obtain crude preparations with stable lipase activity. Using these preparations, optimal conditions for assaying mesocarp lipase activity were determined and further used to identify oil palm breeding lines with low lipase levels producing oils with low acid values (Ngando Ebongue et al., 2008). The gene coding for this lipase was identified in the genome of Elaeis guineenses (Morcillo et al., 2013). It shares a high homology with the lipase from Carica papaya latex (Dhouib et al., 2011) and more generally with fungal lipases of the lipase 3 family, exhibiting around 30 % sequence coverage and 30 % amino acid identity (data not shown). Because of the similarity between oil palm and licuri, it is tempting to search for a similar lipase in licuri.
Lipases from vegetable sources are often referred to as naturally immobilized or self-immobilized, biocatalysts because they can be trapped in the vegetable matrix. As such, they can be applied directly to the reaction medium as biocatalysts (Caro et al., 2000; Villeneuve, 2003). This phenomenon is exemplified by the lipase isolated from Carica papaya latex, which is believed to be associated with polyisoprene molecules within the latex matrix (Abdelkafi et al., 2011; Avelar et al., 2013). Using naturally immobilized plant lipase could be more economical and sustainable when compared to microbial and animal lipases, which require lengthy fermentation and purification processes, as well as immobilization, prior to application (Abdelkafi et al., 2009; Avelar et al., 2013).
Thus, assuming that a lipase is found in Licuri, both the lipase and lipids could be considered for licuri fruit valorization and new bioprocesses based on the concept of biorefinery and circular economy could be developed. Here, we describe our attemps to isolate and characterize a lipase activity present in the mesocarp of Licuri fruit. Since some lipolysis of endogenous lipids was observed, the lipid composition of both the kernel and mesocarp was also investigated.
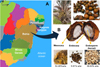 |
Fig 1 Geographical distribution of the licuri palm tree (Syagrus coronata (Mart.) Becc.) and anatomy of its fruit. Licuri is endemic to regions from northern Minas Gerais to southern Pernambuco (panel A). Panel B shows the standard shape and color of the licuri fruit used in this study, along with anatomical features, cross-section, and physical characteristics of ripe, freeze-dried licuri fruits collected from Tingui Farm in Malhada dos Bois, Sergipe, Brazil (Latitude: 10°20′58″ S; Longitude: 36°55′27″ W). |
2 Materials and methods
2.1 Material and reagents
The licuri fruit (Syagrus coronata (Mart.) Becc) was kindly provided by Cyro Rocha da Silva, owner of the Tingui farm, located in Malhada Dos Bois–SE (Latitude: 10° 20' 58" South; Longitude: 36° 55' 27" West). Ripe licuri fruits were selected as samples by having a yellowish and uniform color, without lesions, with a longitudinal diameter of approximately 4.5 cm. The anatomical features of licuri fruit were measured in relation to the total mass of the fruit, comprising the mesocarp, endocarp, and kernel, as described in Figure 1.
The selected fruit samples were sterilized by immersion in 1 % sodium hypochlorite solution for 10 min and then thoroughly washed with distilled water. The fruits were dried by lyophilization and stored in vacuum bags at −20°C. Since oil degradation can occur rapidly after harvest, this step was employed to preserve the whole fruit and ensure that all tests were performed on samples in a comparable state.
The two most prominent fruit fractions, the kernel and mesocarp, were selected and separated to extract lipids and search for lipase activity on tissue homogenates and after delipidation. Since licuri is a native Brazilian palm tree, this work was registered in the National System for the Management of Genetic Heritage and Associated Traditional Knowledge (SisGen) under n° A6D8A4D, in compliance with the provisions of law and its regulations.
The solvents and reagents used were hexane P.A., 99 % (Synth, Brazil); sodium hypochlorite 2.5 % (Miala); ethanol P.A. 99.8 % (Neon); pure gum arabic powder (Synth, Brazil); acetone P.A. 99.5 % (Dinâmica Química Contemporânea Ltda.); potassium hydroxide P.A. 85 % (Dinâmica Química Contemporânea Ltda.); commercial extra virgin olive oil with low acidity (Carbonell); and commercial licuri kernel oil (COOPES). Thin-layer silica gel plates (10 × 20 cm) were from Merck Sigma–Aldrich. The lipid standards monogalactosyldiacylglycerol (MGDG) and digalactosyldiacylglycerol (DGDG) were purchased from Avanti Polar Lipids, while oleic acid, triolein, diolein, and monoolein, used as standards for free fatty acids (FFA), triacylglycerols (TAG), diacylglycerols (DAG), monoacylglycerols (MAG), tributyrin, and the bile salt sodium taurodeoxycholate (NaTDC) were purchased from Sigma–Aldrich. All other analytical-grade reagents were purchased from Synth®, Vetec Química (São Paulo, Brazil), Fisher Scientific, and Merck.
2.2 Dry matter and extraction yield
Although the entire fruits had already been freeze-dried before storage, the drying appeared to be not optimum with this type of fruit having a hard endocarp protecting the kernel. Therefore, frozen kernels were ground in a mill and dried again overnight in an oven at 100°C until a constant weight was achieved and dry matter contents were determined. The whole processed material mass (W2, g) and the separated fractions (kernel and mesocarp) were weighed on a dry basis. The material extraction yield for each fraction (recovered mass W1, g) was estimated from Equation (1).
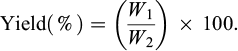 (1)
(1)
2.3 Delipidation of licuri’s kernel and mesocarp
2.3.1 Soxhlet extraction
Lipid extraction of the dry and grounded licuri kernel was carried out in a Soxhlet extractor (Nova Ética), using hexane as a solvent and varying the extraction time from 2 to 8 h, according to the methodology described by Kouteu et al. (2017), with some modifications. The resulting delipidated cakes were dried by vacuum filtration for 1 h and then kept for an additional period of 24 h at 4°C to ensure complete evaporation of hexane. The dilapidated cake samples were freeze-dried and sieved to standardize the particle size below 0.5 mm diameter (using a 35-mesh sieve) and stored at −20°C to be used for further analysis. The kernel oil recovered from the hexane phase was stored at room temperature in amber glass bottles in a location protected from light.
2.3.2 Cold hexane extraction
The delipidation using cold hexane was adapted from previous studies (Pierozan et al., 2009; Ngando Ebongue et al., 2006; Santos et al., 2013) with slight modifications. Frozen licuri kernel and mesocarp were separately soaked in cold hexane 1:5 (w/v) and grounded in a blender for 10 min. Each material was vacuum filtered, and cold hexane was added again at a 1:5 (w/v) ratio. Delipidation was performed under stirring for 1 to 8 h for the kernel, and only 3 h for the mesocarp, due to its lower lipid content. To ensure a maximum removal of hexane, the delipidated cakes were dried by vacuum filtration for 1 h and then kept for 24 h at 4°C. They were then freeze-dried and stored at −20°C until lipase separation and activity measurements. The sample powders were named dry delipidated mesocarp (DDM) and dry delipidated kernel (DDK). The solvent was then separated from the oil by rotary evaporation, and the oil was stored at 4°C in amber glass bottles for subsequent use.
2.4 Fractionation of the dry delipidated licuri mesocarp powder
The dry delipidated mesocarp fraction (DDM; 2.5 g) was resuspended in 20 mL buffer solution (Tris–Base 50 mM (3 g/L−), Triton X-100 0.5 % (v/v−), pH 8) and placed under vigorous stirring at 4⁰C for 24 h. The suspension was then centrifuged at 4000 rpm, 4⁰C for 30 min to facilitate phase separation. After centrifugation, three phases were obtained (liquid, gelatinous interphase, and solid), and lipase activity was measured in all three fractions. Since both the liquid and gelatinous phases contained significant lipase activity, they were pooled and then lyophilized for 24 h. The freeze-drying process was completed by gently grinding the sample in a mortar after immersion in liquid nitrogen. A fine powder was thus obtained and named thereafter pre-purified licuri lipase (PPLL). The entire preparation process, from delipidation to pre-purification, is illustrated in Figure 2.
 |
Fig 2 Diagram of the preparation process for the preliminary characterization of licuri lipase. (A) Preparation of dry delipidated mesocarp (DDM). (B) Preparation of pre-purified licuri lipase (PPLL), including freeze-drying of pooled liquid and gelatinous phases. |
2.5 pH-stat assay of lipolytic activities
Lipase activities of mesocarp homogenate, DDM, DDK, and PPLL were assayed potentiometrically by automatically titrating the free fatty acids (FFAs) released from mechanically stirred triglyceride emulsions in a thermostated (37°C) vessel, using 0.1 N NaOH and a pH-stat device (718 Stat Titrino, Metrohm) as described by Ngando-Ebongue et al. (2006). Various pH endpoints ranging from 4 to 10 were tested with each substrate. For the assay with short-chain triglycerides (tributyrin; TC4), the reaction medium had the following composition: 3.33 % (v/v) TC4, 0.5 to 60 mM NaTDC, 7.5 mM CaCl2, 0.33 mM Tris–HCl, and 150 mM NaCl. TC4 was added directly to the pH-stat vessel containing the assay solution and was emulsified by mechanical stirring using a propeller. The total reaction volume ranged from 15 to 30 mL depending on the reaction vessel that was used.
Medium-chain triglycerides (licuri oil; mainly C12 fatty acids) or long-chain triglycerides (olive oil; mainly C18:1) were first pre-emulsified with gum arabic (GA) by mixing 5 mL of oil with 45 mL of a 10 % (w/v) GA solution prepared in 7.5 mM CaCl2, 0.33 mM Tris–HCl, and 150 mM NaCl. Five milliliters of the oil–GA emulsion were then used as a substrate and mixed in the pH-stat vessel with 10 ml of the assay solution. Final concentrations in the reaction medium were 3.33 % (v/v)− oil, 7.5 mM CaCl2, 0.33 mM Tris–HCl, 150 mM NaCl, and 3.3 % (w/v)− GA in routine assays. In some assays, various concentrations of GA were tested.
The pH-stat technique was also used to assay phospholipase activity using egg L-α-phosphatidylcholine (Egg PC) (Sigma) as a substrate, at 37°C and pH 8.0 according to Amara et al. (2012), as well as the esterase and lipase activities on vinyl propionate according to Chahinian et al. (2002).
For each assay, 1 unit (U) of lipase activity corresponds to 1 μmol of fatty acid released per minute under the assay conditions used.
2.6 Assay of protein contents, electrophoresis, in-gel tryptic digestion, and protein identification by mass spectrometry
Protein content was determined by using the Pierce BCA Protein Assay Kit (Pierce Biotechnology, Rockford, IL, USA) with bovine serum albumin as a reference standard. Electrophoresis of protein samples was done using 12 % (w/v−) SDS–PAGE, and the gels were stained with Coomassie Brilliant Blue R-250. The most intense bands were cut and submitted to trypsin digestion as previously described by Abdelkafi et al. (2012). LC-MS/MS analyses were performed on an ESI-Q-Exactive plus mass spectrometer (ThermoFisher) coupled to a nano liquid chromatography (Ultimate 3000, Dionex) as described by Zhang et al. (2018).
For protein identification, the mass spectra were processed using the Proteome Discoverer software (Thermo Fisher, version 2.1.0.81), with the NCBI database of the following species: Carica papaya (TaxID: 3649), Cocoseae (TaxID: 169705), Elaeis guineensis (TaxID: 51953), Phoenix dactylifera (TaxID: 42345), Syagrus coronata (TaxID: 682621), Arabidopsis thaliana (TaxID: 3702), and Thermomyces lanuginosus (TaxID: 5541).
2.7 Lipid analysis
2.7.1 Thin-layer chromatography
Thin-layer chromatography (TLC) analysis of the lipid extracts in hexane was performed according to Wattanakul et al. (2019) with some modifications. Lipid extract samples and reference standards (DGDG, MGDG, TAG, FFA, DAG, and MAG) were spotted as a 5 mm band onto a thin-layer silica plate using a Linomat IV sample spotter (Camag) equipped with a 100 μL Hamilton syringe. Elutions were performed with chloroform/methanol/water (25/15/1.25, v/v/v) for polar lipids and heptane/ethyl ether/formic acid (55/45/1, v/v/v) for the separation of neutral lipids and free fatty acids.
After elution, the plates were dried at room temperature in a fume hood. Thymol-sulfuric acid and copper acetate-phosphoric acid solutions were previously prepared for the staining of polar lipids (galactolipids) and neutral lipids, respectively. The dry plates were submerged in these solutions using a Chromatogram Immersion Device 3 (Camag, Muttenz, Switzerland) and then subsequently dried again in a fume hood at room temperature to evaporate the excess staining solution before being heated in an oven at 100°C for 10 min (polar lipids) or at 180°C for 15 min (neutral lipids). Pictures of the plates were taken, and the separate compounds were identified by comparison with the standards used. Neutral lipids were quantified using calibration curves established with triolein, oleic acid, diolein, and monoolein as reference standards (see Fig. S1).
2.7.2 Fatty acid analysis by gas chromatography with flame detector
The analyses were performed at the Multiuser Chemistry Laboratory Center (CLQM), located in the Department of Chemistry—UFS. After hydrolysis and derivation of lipids into fatty acid methyl esters (FAME) according to Barbosa et al. (2019), the FAME separation was performed on a Shimadzu gas chromatograph, model 2010 PLUS, equipped with a flame ionization detector and a capillary Supelcowax 10 column (30 m × 0.25 mm ID, 0.25 μm film thickness). The carrier gas was helium (He) at a constant flow rate of 1.2 mL/min. For the FID detector, the gas flows were make-up gas (N2) at 30 mL/min, hydrogen (H2) at 30 mL/min, and synthetic air at 300 mL/min.
The program used for the temperature ramp was 80°C (1 min), ramped at 10°C/min to 210°C (1 min), then ramped at 5°C/min to 230°C and held for 10 min. A sample volume of 1 μL was automatically injected in split mode (1:20), SCAN mode, with linear velocity mode, detector and injector temperatures of 230 °C, and a total analysis time of 29 min.
3 Results and discussion
3.1 Characterization of lipid extracts from licuri kernel and mesocarp
Lipid extracts containing both polar lipids (mainly galactolipids) and neutral lipids (predominantly triglycerides) were obtained from licuri kernel and mesocarp using hexane under various conditions.
For the licuri kernel, the highest lipid extraction yield (50.8 ± 3.6 % w/w) was achieved using Soxhlet extraction for 8 h after oven-drying the kernels (Table 1). A shorter extraction time of 6 h yielded a comparable result (48.4 ± 4.4 % w/w), indicating that prolonged extraction had only a marginal effect on total lipid recovery. These values are consistent with previously reported lipid contents for Licuri kernels (49.2 % w/w) (Crepaldi et al., 2001), confirming the reliability of the extraction methodology applied in this study.
Drying in a desiccator yielded lower values than drying in an oven (Table 1). This variation in extraction yield could result from higher moisture levels in the powder (Farzaneh et al., 2017). It was effectively checked that the oven and dessicator drying procedures gave moisture percentages of 0.6 % and 8 % w/w, respectively.
When lipid extraction was performed by grinding the licuri kernel in cold hexane, it was possible to extract 30.3 % w/w of lipids. An additional stirring period of 3 h at 4°C allowed reaching a lipid extraction yield of 40.1 ± 2.9 % w/w, i.e., 50.8 % of the maximum extraction yield obtained with Soxhlet after 8 h of extraction (Table 1).
In line with our findings, previous studies reported similar licuri kernel oil yields—49.2 % w/w (Crepaldi et al., 2001) and 50 % w/w (Iha et al., 2014)—even though fruits were harvested in distinct Brazilian states (Bahia and Alagoas, respectively). By contrast, a lower yield of 39 % w/w was observed by De la Salles et al. (2010).
Since the cold hexane extraction allowed a partial delipidation of around 80 %, the level that allowed keeping the lipase activity stable in the previous work of Ngando-Ebongue on oil palm fruit (Ngando Ebongue et al., 2006), this procedure was chosen for the delipidation of licuri mesocarp. It allowed reaching a lipid extraction yield of 11.2 % w/w for 3 h of stirring (Table 1). Previous works on licuri mesocarp by Crepaldi et al. (2001) and Xavier et al. (2015) reported a lower lipid extraction yield of 4 % w/w, although they used the Soxhlet extraction method. This discrepancy might result from the region where the licuri fruit was harvested, as well as the level of maturation.
The cold hexane lipid extracts obtained from licuri kernel and mesocarp after 3 h were then analyzed by TLC, as shown in Figure 3. Concerning licuri kernel neutral lipids (Fig. 3A), a major but diffused band corresponding to TAG was observed, which migrated slightly below the triolein reference standard. This probably reflected the high contents of saturated medium-chain fatty acids in the TAG from licuri kernel oil (Iha et al., 2014) (also see Table 2). Regarding polar lipids in licuri kernels, no significant band was observed. (Fig. 3B).
In the case of mesocarp neutral lipids (Fig. 3A), the band corresponding to TAG was extremely weak, but an intense band corresponding to FFA was observed, indicating that TAG degradation by an endogenous lipase occurred. Similarly, the analysis of polar lipids and staining with thymol-sulfuric acid (Fig. 3B) revealed the presence of bands that could correspond to galactolipid lipolysis products like digalactosyl monoacylglycerol (DGMG), while no bands corresponding to MGDG and DGDG were observed. An upper band corresponding to FFA was also observed (Fig. 3B). The analysis of mesocarp oil by Fourier-transform infrared spectroscopy also revealed a shift toward lower wavenumbers for the stretching vibration of carbonyl groups, indicating the presence of FFA (see Fig. S6 in Supporting Information). Further analysis of polar lipids by TLC and staining with the molybdenum blue reaction did not reveal the presence of phospholipids (data not shown). We assumed that extraction with hexane was not efficient for the recovery of these lipids.
Figure 3C shows the conversion of MGDG and DGDG reference standards into MGMG and DGMG using pancreatic lipase-related protein 2 (PLRP2) as previously studied by (Amara et al., 2010). By comparison, the presence of DGMG in the lipid extract of licuri mesocarp (Fig. 3B) suggests the presence of an endogenous galactolipase activity in licuri mesocarp. This hypothesis was confirmed after blanching licuri mesocarp (5 min in boiling water) prior to lipid extraction. Blanching results in the heat inactivation of endogenous enzymes (Jha et al., 2013; Wattanakul et al., 2019), allowing for the visualization of a band corresponding to DGDG that was not present in the non-blanched mesocarp lipid extract (Fig. 4A). Staining for neutral lipids (Fig. 4B) also revealed that blanching allowed decreasing the intensity of the band corresponding to FFAs. These findings confirmed the presence of endogenous lipolytic activities in the mesocarp of licuri.
Another result that reflects the presence of lipolytic activities in the licuri mesocarp is that after blanching, a higher lipid extraction yield of 13.1 % w/w was obtained compared to 11.2 % w/w with non-blanched mesocarp. These values are higher than the lipid contents previously reported (4.5 % w/w) for the Licuri fruit mesocarp (Crepaldi et al., 2001). Quantitative TLC analysis of neutral lipids from non-blanched mesocarp allowed estimating an FFA level of 9 % w/w (Fig. S1), indicating that 80 %°of lipids were hydrolyzed.
The total lipids extracted from licuri kernel and mesocarp were further analyzed for their fatty acid contents after derivation into FAMEs (Table 2). As expected from the previous literature (Iha et al., 2014), the lipids from the licuri kernel, mainly triglycerides, were rich in saturated medium-chain fatty acids (C8–C12; 67 % w/w), the main fatty acid, lauric acid, accounting for 49.0 ± 0.3 % w/w of total fatty acids. With that respect, licuri oil appears highly similar to palm oil, also obtained from kernels (Table 2; Sehgal and Sharma, 2020). Unlike what is found in licuri kernel, total lipids from licuri mesocarp contain longer and unsaturated fatty acids like palmitic acid (C16:0; 41.5 ± 0.4 %), followed by oleic acid (C18:1; 34.6 ± 0.1 %) and linoleic acid (C18:2; 20.8 ± 0.2 %), thus having a composition closer to that of other vegetable oils. This is the first report to our knowledge of licuri mesocarp fatty acid composition, and it also appears similar to what was reported for oil palm mesocarp (Sehgal and Sharma, 2020). These results could give more added value to licuri fruit as a source of two distinct oils with different applications. Indeed, the mesocarp oil rich in unsaturated fatty acids (C18:1 and C18:2 n-6) could be more interesting for human nutrition and the prevention of cardiovascular diseases, diabetes, and cancer (Huerta-Yépez et al., 2016) than the kernel oil, this latter presenting interesting properties for cosmetics and lipid-based pharmaceutical formulation (Carrière, 2016). Moreover, its high content of medium chain fatty acids presents an interest for making biofuels either by transesterification (De La Salles et al., 2010) or by photo-enzymatic conversion of fatty acids into alkanes (Sorigué et al., 2017).
Considering the results on lipid characterization presented in this section, we can conclude that licuri mesocarp oil shows great potential due to its favorable fatty acid composition. However, efforts must be made to reduce the levels of FFA, as oils with FFA levels above 5 % are considered unsuitable for human consumption (Ngando Ebongue et al., 2008). Therefore, controlling the catalytic activity of the endogenous lipase is essential to improve oil quality. In this context, the following section aims to characterize and better understand the catalytic activity of the lipase presumably present in licuri mesocarp.
Lipid extraction yields (% w/w) from Licuri kernel and mesocarp using hexane in a Soxhlet apparatus (with prior drying in oven or desiccator) or cold hexane. Values are mean ± SD (n = 3) and are expressed on a dry basis.
 |
Fig 3 Thin-layer chromatography analysis of lipids extracted from the Licuri mesocarp (Mes) and endocarp (End) with cold hexane. (A) Separation of neutral lipids stained with copper acetate-phosphoric acid. DAG, Diacylglycerol (Rf: 1.3-DAG: 0.31 ± 0.02; 1.2(2.3)-DAG: 0.25 ± 0.02); End, endocarp or kernel; FFA, Free fatty acids (Rf: 0.48 ± 0.03); MAG, Monoacylglycerol (Rf: 0.02 ± 0.01); Mes, Mesocarp; Rs, Reference standards including triolein, oleic acid, diolein, and monoolein; TAG, Triacylglycerol (Rf: 0.88 ± 0.05). (B) Separation of polar lipids stained with thymol-sulfuric acid. End, endocarp; MGDG: monogalactosyl diacylglycerol (Rf: 0.63 ± 0.05); DGDG: digalactosyl diacylglycerol (Rf: 0.31 ± 0.02). (C) Hydrolysis of MGDG and DGDG reference standards by pancreatic lipase-related protein 2 (PLRP2), leading to the formation of monogalactosyl monoacylglycerol (MGMG; Rf: 0.40 ± 0.03) and digalactosyl monoacylglycerol (DGMG; Rf: 0.10 ± 0.02). The samples were obtained according to Sahaka et al. (2021). Original pictures are available in the Supporting Information (Figs. S1, S2 and S3). |
Comparison of fatty acid composition (relative% w/w) of kernel and mesocarp of licuri and oil palm fruits. afrom (Iha et al., 2014); bfrom (Sehgal and Sharma, 2021).
 |
Fig 4 Thin-layer chromatography analysis of lipid extracts from licuri mesocarp obtained with and without blanching. (A) Polar lipids stained with thymol-sulfuric acid. (Rf: FFA: 0.69 ± 0.05; MGDG: 0.63 ± 0.05; MGMG: 0.40 ± 0.03; DGDG: 0.31 ± 0.02; DGMG: 0.10 ± 0.02). (B) Non-polar lipids stained with copper acetate-phosphoric acid. FFA (Rf: 0.62 ± 0.05); MGDG: monogalactosyl diacylglycerol (Rf: 0.54 ± 0.05); DGDG: digalactosyl diacylglycerol (Rf: 0.29± 0.02). Original pictures are available at Supporting information (Figs. S4 and S5). |
3.2 Search for lipase activity in the licuri mesocarp and kernel
Freeze-dried licuri kernel and mesocarp were homogenized in either 50 mM Tris–HCl, 0.5 % Triton X-100, pH 8, or 10 mM MES, 0.5 % Triton X-100, pH 6, buffers (3.5 g in 20 mL) using an Ultra-Turax. Lipase activity in these homogenates was measured using tributyrin (TC4) as a substrate at pH 7. These homogenates were then kept at 4°C for one month, and the variation in their lipase activity was measured. While no significant activity was recorded for the kernel homogenate, the licuri homogenate showed a lipase activity of 283.73 ± 17.61 U per g of freeze-dried mesocarp at pH 6 and 227.77 ± 14.13 U per g at pH 8. These activities were then found to decrease with time at 4°C, reaching a residual activity of 25 % after one month (Fig. 5A). Apart from the initial value that was higher for pH 6, no significant differences were observed between the two pH levels of homogenization. The presence of a much higher lipase activity in the mesocarp than in the kernel is in good agreement with the endogeneous lipolysis of lipids only observed in mesocarp (Figs. 3 and 4).
It is worth noting that similar levels of lipase activity were previously found in homogenates of fresh oil palm mesocarp (190 ± 17 U/g). Since the lipase activity present in total mesocarp homogenate appeared to be unstable with time, even at 4°C, we then proceeded to a delipidation of tissues using cold hexane, a procedure that previously led to a more stable lipase preparation from oil palm mesocarp without reducing the enzyme activity (Ngando Ebongue et al., 2006). DDM and DDK were recovered after filtration and freeze-drying (Fig. 2), and then their lipase activities on both tributyrin and olive oil were tested after dispersion of the powder in 50 mM Tris–HCl, 0.5 % Triton X-100, pH 8, buffer (Table 3). Again, lipase activity in the DDK sample obtained from the kernel was very low (<4 U/g). In DDM, the lipase activity on tributyrin was 202.0 ± 13.0 U/g of delipidated powder, a value that represented 88.7 % of the lipase activity initially measured in mesocarp homogenate before delipidation. Thus, delipidation of the mesocarp with cold hexane did not cause deleterious effects to the enzyme. Moreover, the lipase activity of DDM dispersion appeared more stable when kept at 4°C, with a residual activity of 95 % after 21 days (Fig. 5).
To confirm that the observed activity was not obtained through other factors than enzyme activity, a heat inactivation (100°C for 1 h) of the DDM sample was also carried out in a closed container to avoid moisture variation. Activities on both tributyrin and olive oil were suppressed (Table 3).
 |
Fig 5 Stability of licuri mesocarp lipase. (A) Variation with time in the lipase activity of licuri mesocarp homogenates and dry delipidated mesocarp (DDM) dispersion kept at 4°C. (B) Residual activity of PPLL after incubation at either room temperature (with and without protease inhibitors) or 4°C. Lipase activity was measured using tributyrin (TC4) as a substrate at pH 7. |
Lipase activities (U·/g−) in the powders obtained from kernel (DDK) and mesocarp (DDM) from licuri fruit. Activities were measured at pH 7 using either olive oil (3% gum Arabic) or tributyrin as substrates.
3.3 Optimization of lipase activity measurements in dry delipidated licuri mesocarp
To optimize the measurement of lipase activity present in DDM, we tested several parameters, including the triglyceride substrate (olive oil and tributyrin), the pH and the presence of surfactants such as gum arabic and bile salts NaTDC. Lipase activity was expressed either in units (U) per g of DDM powder or in U per mg of proteins after the concentration of proteins in DDM was determined.
Assays with olive oil were initially performed in the presence of 3 % gum Arabic, an emulsifier commonly used for stabilizing vegetable oil emulsions (Tiss et al., 2001). GA is a hetero-arabino-galactan polysaccharide (containing about 2 % of proteins), characterized by a structural flexibility at the oil-water interface due to its contents in hydrophilic sugar and aromatic amino acids (Sanchez and Demain, 2011). Under these conditions, lipase activity was found to be maximum at pH 8 with specific activities of 208.33 ± 7.22 U DDM/g− and 10.7 ± 1.1 U protein/mg−, respectively (Fig. 6A).
This value is in the same range as the lipase activities on olive oil measured with the lipases from dry Carica papaya latex (256 ± 8 U/g− (Abdelkafi et al., 2011)) and oil palm (Elaeis guineenses Jacq.) mesocarp dry powder (250 ± 14 U DDM/g− (Ngando Ebongue et al., 2006)), these two activities showing, however, an optimum at pH 9. A lipase found in coconut kernel (Cocos nucifera linn) had, however, an optimal pH in the 7.5 to 8.5 range (Ejedegba et al., 2007).
The effect of GA concentration was then tested, and the activity was found to increase when GA was decreased, reaching a maximum value of 375 ± 12 U/g− at 1 % GA at pH 8 (Fig. 6B). These findings probably reflect the fact that surfactants can favor lipase activity as emulsifiers but can also inhibit it at high concentrations by competing with the enzyme for interfacial adsorption (Delorme et al., 2011).
A similar effect was observed when testing the effect of GA on the interfacial binding of human pancreatic lipase (Tiss et al., 2001). At a final concentration of 3 % GA, it was possible to observe that human pancreatic lipase was desorbed from the oil-water interface.
Since bile salts are other surfactants commonly used in lipase assays, we also tested the effect of NaTDC concentration on DDM activity using tributyrin as substrate at pH 7 (Fig. 6C). An initial concentration of 0.5 mM NaTDC is often used with this substrate to reduce surface pressure and avoid the interfacial denaturation of lipases (Aloulou et al., 2007). An activity of 229 ± 7 U/g− was measured under these conditions. It then increased up to 475 ± 25 U/g− at 10 mM NaTDC and slightly decreased to 350–400 U/g− at higher concentrations. DDM lipase was therefore active at NaTDC concentrations far above the critical micellar concentration (CMC) of NaTDC (1–4 mM at 20–25°C), showing no evidence of inhibitory effect of bile salts on Licuri lipase.
Only certain types of lipases, such as gastric lipase (Chahinian et al., 2006), Yarrowia lipolytica lipase (Aloulou et al., 2007), and Carica papaya latex lipase (Abdelkafi et al., 2011), are not inhibited by bile salts above their CMC. Therefore, like these lipases, DDM may also be a good candidate for acting in the gastrointestinal tract and be used in pancreatic enzyme replacement therapy.
 |
Fig 6 Effect of several parameters on lipase activity in licuri mesocarp. (A) Variation with pH in the lipase activity on olive oil in the presence of 3% w/v gum arabic. (B) Effect of Gum Arabic (GA) concentration on the hydrolysis of olive oil at pH 8. (C) Effect of bile salts (NaTDC) concentration on tributyrin hydrolysis reaction at pH 7. |
3.4 Fractionation and activity on various substrates of dry delipidated licuri mesocarp
The DDM fraction was used for lipase isolation trials. DDM powder (2.5 g) was resuspended in Tris buffer, pH 8, containing 0.5 % (v/v) Triton X-100 (v/v−) and kept under vigorous stirring at 4⁰C for 24 h. After centrifugation at 4000 rpm, 4⁰C for 30 min, three phases (liquid, gelatinous interphase, and solid pellet) were obtained, and their lipase activity was measured. While the solid pellet was poorly active and discarded, the liquid and gelatinous phases showing significant lipase activity were pooled and lyophilized. The final mass recovery after lyophilization was 56.6 ± 15.8 % of DDM powder, and the lipase activity expressed in U per g of dry powder reached 700 U/g− and was increased three-fold. However, the lipase-specific activity expressed in U per mg of proteins was not increased, indicating that the removed pellet of insoluble material did not consist of proteins. A pre-purified licuri lipase (PPLL) was thus obtained as a powder that could be used in biocatalysis or for further lipase enrichment and purification. PPLL was further characterized for its substrate specificity and its optimum pH of activity using various lipid substrates (Fig. 7A).
PPLL displayed its highest specific activity (17.7 ± 1.3 U/mg protein at pH 5.5) on licuri kernel oil, mostly consisting of medium-chain fatty acids. The second-best substrate was olive oil with a PPLL specific activity of 13.5 ± 0.1 U/mg protein at pH 7.5–8.0. Specific activity on tributyrin was 10.1 ± 0.2 U/mg protein at pH7.0 and therefore, no correlation between the level of activity and the chain length of fatty acids could be established.
PPLL was also found to display some phospholipase activity on egg PC, but this activity was weaker with a maximum value of 1.2 ± 0.2 U/mg protein at pH 8.
Finally, PPLL activity was tested on vinyl propionate, a short-chain ester showing a high solubility in water (S = 86 mM). The highest activity (9.3 ± 0.4 U/mg protein) was recorded at pH 8.5. At higher pH values, some spontaneous hydrolysis of vinyl propionate precluded the measurement of reliable enzyme activity. This substrate was also used to compare the activities of PPLL on soluble and insoluble substrates by using either a concentration of 30 mM (i.e., below the saturation) or 153 mM (i.e., above saturation, with the formation of an oil-in-water emulsion). PPLL was found to be five-fold more active on emulsified than on soluble vinyl propionate (Fig. 7B), suggesting an interfacial activation (Ferrato et al., 1997), a characteristic of true lipases acting preferentially on water-insoluble long-chain TAGs. In contrast, typical esterases are not interfacially activated, and they show their highest activity on water-soluble esters. Short chain vinyl esters are thus convenient substrates to discriminate between lipases and esterases (Chahinian et al., 2002).
From Figure 7, one can clearly see that PPLL has the capacity to hydrolyze a variety of lipid substrates, with a preference for medium- and long-chain TAG and thus exhibits the behavior of a true lipase. The observation of different pH optima depending on the substrate suggests that PPLL may be a mixture of several lipolytic enzymes. However, it is known that single lipases can display different pH optima depending on the TAG substrate, which can be explained by the pH-dependency of the interfacial adsorption that varies with the type of TAG and the interfacial properties (Chahinian et al., 2006).
Similarly, the detection of a phospholipase activity of PPLL on egg PC does not necessarily mean the presence of both phospholipase and lipase since some lipases are well known to display phospholipase A1 activity (Aloulou et al., 2006).
 |
Fig 7 Specific activity of pre-purified licuri lipase extract (PPLL) on various lipid substrates. (A) pH-dependent activities on olive oil, licuri oils tributyrin, phospholipids (Egg PC), and vinyl ester. Vinyl propionate was used at a concentration below its solubility in water (<86 mM; 100 μL/30 mL per assay) to measure the esterase activity of licuri lipase on a soluble substrate. (B) Interfacial activation: PPLL activity on vinyl propionate at substrate concentrations below (30 mM) and above (153 mM) the saturation (86 mM). |
3.5 PPLL storage stability
The stability of PPLL was also tested and stored under different conditions (Fig. 5B). It was observed that PPLL lost half of its initial activity after 10 days of storage at room temperature (RT), becoming completely inactivated after 21 days. In the presence of protease inhibitors, a slight improvement in stability was noted, with 20.7 % of the initial activity still retained after 21 days at RT. Storage at 4°C markedly improved PPLL stability, with 75.4 % of its initial activity maintained after one month. This stability of PPLL was similar to that of DDM kept for one year at −20°C (data not shown).
Other plant lipase preparations demonstrated similar behavior, such as castor bean (Ricinus communis L.) lipase that was kept at 4°C for seven months and retained between 80 % and 90 % of its initial activity (Cavalcanti et al., 2007).
3.6 Preliminary SDS–PAGE and proteomics analysis of PPLL
To obtain more information on the proteins present in the PPLL preparation, SDS–PAGE analysis was performed (Fig. 8).
Several bands are observed, with the predominance of three bands of around 25 kDa and the presence of faint bands at 20 kDa and in between 30 and 85 kDa. Plant lipases with molecular masses in this range were previously described in the literature, such as Carica papaya (50–55 kDa (Rivera et al., 2017), Elaeis guineensis (55 kDa; (Morcillo et al., 2013) and in Ricinus communis (60 kDa; (Cavalcanti et al., 2007) lipases. In addition, lipases with lower molecular masses were also described in Heliantus annuus (22 kDa; (Sagiroglu and Arabaci, 2005)). All the visible bands on SDS–PAGE were excised and subsequently submitted to trypsin digestion and proteomics by LC-MS/MS.
The most relevant matches were related to plant GDSL esterase/lipase and phospholipase D (Table 4). Interestingly, the same enzymes were previously detected and identified in Carica papaya latex (Abdelkafi et al., 2012; Abdelkafi et al., 2009), while the detection of the main lipase of interest was more difficult and required the use of a covalently bound radiolabeled lipase inhibitor (Dhouib et al., 2011). Other proteins from Syagrus coronata were identified based on the known genome of Syagrus coronata chloroplast (Áquila et al., 2018), such as ATP synthase CF1 α-subunit (accession number: ALT55379.1; 14 % protein coverage with 5 peptides; 514 amino acids; 56 kDa),ATP synthase CF1 beta chain (accession number: YP_009229388.1; 5 % protein coverage with 2 peptides; 498 amino acids; 53.7 kDa), photosystem II protein D1 (accession number: ALT55393.1; 6 % protein coverage with 2 peptides; 353 amino acids; 38.8 kDa), and ribulose bisphosphate carboxylase (RuBisCO) large chain (accession number: ALT55385.1; 4 % protein coverage with 2 peptides; 486 amino acids; 53.9 kDa). GDSL esterase/lipase and phospholipase D of Syagrus coronata were not found, however, to be encoded within the chloroplast genome, and one will have to wait for the complete genome sequencing or mRNA isolation to obtain the complete protein sequences. No peptide homologous to known lipases from Carica papaya and Elaeis guineensis could be identified, suggesting that in Syagrus coronata, the GDSL esterase/lipase could be the enzyme displaying the lipase activity we measured on various substrates. However, a previous study in Carica papaya has shown that the homologous GDSL enzyme found in this species displays a high esterase activity on vinyl esters and a weak lipase activity on medium and long chain TAGs (Abdelkafi et al., 2009). Therefore, we do not trust that the lipase activity measured in Syagrus coronata mesocarp results from GDSL.
The large band observed at the level of the stacking gel (Fig. 8) suggests that some material was retained there, which could correspond to proteins/enzymes covalently bound to solid particles. Carica papaya lipase is known, for instance, to be naturally immobilized on particles that have so far not been characterized yet (Abdelkafi et al., 2011).
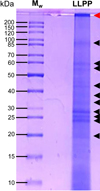 |
Fig 8 SDS–PAGE analysis of PPLL. Black arrows indicate the various protein bands present in LLPP. The red arrow indicates the material remaining on the stacking gel. |
Protein matches with high similarity to lipolytic enzymes obtained from the peptide mass fingerprint of PPLL protein bands separated by SDS-PAGE. Species names: Cocos Nucífera, coconut; Elaeis guineensis, oil palm; Carica papaya, papaya; Phoenix dactylifera, date palm. PSM, number of peptide single matches.
4 Conclusion
This study demonstrates, for the first time, that the licuri mesocarp—an undervalued, high-mass fraction of the fruit—can serve both as a source of unsaturated oil and as a self-immobilized, highly active biocatalyst. Cold-hexane extraction yielded 11.2 % (w/w) mesocarp oil, increased to 13.1 % (w/w) by blanching, which also reduced endogenous lipolysis and improved oil quality. In contrast to the lauric-rich kernel oil (≈40 % yield; ≈50 % lauric acid), mesocarp oil was richer in long-chain fatty acids (palmitic 41.5 %, oleic 34.6 %, and linoleic 20.8 %), aligning with oil palm—like profiles. DDM exhibited high lipase activity (375 ± 12 U/g on olive oil, pH 8; 475 ± 25 U/g on tributyrin, pH 7). These activities can appear as low compared to the specific activities of some purified lipases, reaching several thousand U/mg of protein. Nevertheless, these activities are among the highest lipase activities found in crude plant tissues (Abdelkafi et al., 2011). Considering that 1.8 g of DDM can be obtained from a lyophilized licuri fruit weighing on average 5.8 g, i.e., 31 % w/w, and that DDM contains a lipase activity of 375 ± 12 U/g on olive oil, one can thus estimate that 1 kg of licuri fruit contains 116250 (olive oil) lipase units. For comparison, pancreatin from porcine pancreas, a common natural source of lipase, contains a lipase activity of ≥2 U/mg on olive oil (pH 9, 37°C) according to the US Pharmacopeia (USP, 2018)), i.e., ≥200000 U/kg. The whole licuri fruit thus contains a lower lipase activity compared with pancreatin, but DDM appears as a competitive product with 1.9-fold the minimum activity recommended by USP for pancreatin.
Partial solubilization with 0.5 % Triton X-100 followed by phase pooling and lyophilization produced a preparation (PPLL) enriched ∼2-fold (≈700 U/g on tributyrin, pH 7) with the highest specific activities on licuri kernel oil (17.7 ± 1.3 U/mg protein, pH 5.5) and olive oil (13.5 ± 0.1 U/mg, pH 7.5–8.0). Activity on insoluble TAGs and interfacial activation on vinyl esters indicate a true lipase. DDM retained activity over long-term freezing (–20°C, 1 year), and PPLL remained active under short-term refrigeration (4°C, 1 month).
Proteomics detected a phospholipase D and a GDSL esterase/lipase in PPLL, but no canonical lipase peptides, suggesting that the major lipase is in low abundance and/or particle bound, hampering standard SDS–PAGE/LC–MS identification. Future work should pursue activity-guided purification (e.g., mild aqueous biphasic systems and interface-focused fractionation), covalent activity probes, and optimized peptide mapping to pinpoint the catalytic entity.
Overall, valorizing mesocarp oil and the PPLL biocatalyst supports an integrated licuri biorefinery vision—generating added-value products (lauric, oleic, and linoleic-based streams; biocatalyst for lipid transformations) and reducing waste within a circular-economy framework. In this context, Licuri becomes a strategic feedstock for sustainable development in Brazil's semiarid regions, aligning environmental conservation with social and economic growth through circular bioeconomy principles.
Acknowledgements
We are grateful to Dr Régine Lebrun and the Proteomics platform from Institut de Microbiologie de la Méditerranée (IMM, FR3479), Marseille, France.
Funding
The authors are grateful for the financial support by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES, Finance Code 001), specially for the scholarship grants provided for Milena Chagas Lisboa (PROSUP - 88882.365551/2019-01, PDSE 41/2018 - 88881.361582/2019-01).
Conflicts of interest
The authors declare no existing conflict of interest.
Author contribution statement
Milena Chagas Lisboa: Investigation–, Formal Analysis, –Data Curation, Funding Acquisition, Writing – Original Draft Preparation, Writing – Review & Editing; César de Almeida Rodrigues: Investigation–, Formal Analysis, Writing – Original Draft Preparation, Writing – Review & Editing; Moulay Sahaka: Methodology–, Investigation, Formal Analysis; Hélène Launay: Funding Acquisition, Writing – Review & Editing; Ranyere Lucena de Souza: Writing – Review & Editing; Frédéric Carrière: Conceptualization–, Supervision–, Formal Analysis, Methodology–, Writing – Review & Editing; Álvaro Silva Lima: Formal Analysis–, Methodology–, Writing – Review & Editing; Cleide Mara Faria Soares: Supervision–, Funding Acquisition, Resources, Formal Analysis, Methodology, Writing – Review & Editing.
Supplementary Material
Fig. S1. Original picture for Figure 2A. Left: Endosperm (Kernel) runs, Middle: Calibration curves with TAG + FFA + DAG + MAG; Right: Mesocarp. Used in edition: 1 lane from endosperm (kernel), 1 lane from mesocarp, and one lane of the calibration curve.
Fig. S2. Original Picture for Figure 2B. Left: Kernel runs; Middle: Standards (MGDG and DGDG); Right: Mesocarp runs. Used in edition: 1 lane kernel, 2 lanes standard and 1 lane mesocarp.
Fig. S3. Original picture for Figure 2C. TLC analysis of reference standards (MGDG, lane 1 from the left; DGDG, lane 2) before and after incubation with pancreatic lipase-related protein 2 (PLRP2), lanes 2 and 4, respectively.
Fig. S4. Original picture for Figure 3A. Left: Mesocarp without blanching (50 μL, 50 μL, 100 μL, 100 μL); Middle: Standards (MGDG and DGDG); Right: Mesocarp with blanching (50 μL, 50 μL, 100 μL, 100 μL). Used in edition: Standards, 1 lane with, 1 lane without blanching.
Fig. S5. Original picture for Figure 3B. Left: Mesocarp without blanching (50 μL, 50 μL, 100 μL, 100 μL); Middle: Standards (MGDG and DGDG); Right: Mesocarp with blanching (50 μL, 50 μL, 100 μL, 100 μL). Used in edition: Standards, 1 lane with, 1 lane without blanching.
Fig. S6. FTIR spectrum of the fractions obtained from the lipid extracts of licuri kernel oil (A) and mesocarp (B). All fractions obtained in the process were previously lyophilized and analyzed by Fourier transform infrared spectroscopy (FTIR) in an Agilent Cary 630 FTIR spectrometer at 25°C, using the transmission method in the wavenumber range between 4000 and 500 cm−1, with 32 scans and a resolution of 4 cm−1 in triplicate.
Access Supplementary MaterialReferences
- Abdelkafi S, Barouh N, Fouquet B, et al. 2011. Carica papaya lipase: a naturally immobilized enzyme with interesting biochemical properties. Plant Foods Hum Nutr 66: 34–40. [Google Scholar]
- Abdelkafi S, Abousalham A, Fendri I, et al. 2012. Identification of a new phospholipase D in Carica papaya latex. Gene 499: 243–49. [Google Scholar]
- Abdelkafi S, Ogata H, Barouh N, et al. 2009. Identification and biochemical characterization of a GDSL-motif carboxylester hydrolase from Carica papaya latex. Biochim Biophys Acta Mol Cell Biol Lipids 1791: 1048–56. [Google Scholar]
- Abdelmoez W, Mustafa A. 2014. Oleochemical industry future through biotechnology. J Oleo Sci 63: 545–54. [Google Scholar]
- Aloulou A, Puccinelli D, De Caro A, et al. 2007. A comparative study on two fungal lipases from Thermomyces lanuginosus and Yarrowia lipolytica shows the combined effects of detergents and pH on lipase adsorption and activity. Biochim Biophys Acta Mol Cell Biol Lipids 1771: 1446–56. [Google Scholar]
- Aloulou A, Rodriguez JA, Fernandez S, et al. 2006. Exploring the specific features of interfacial enzymology based on lipase studies. Biochim Biophys Acta Mol Cell Biol Lipids 1761: 995–1013. [Google Scholar]
- Amara S, Barouh N, Lecomte J, Lafont D, Robert S, Villeneuve P, De Caro A and Carrière F. (2010). Lipolysis of natural long chain and synthetic medium chain galactolipids by pancreatic lipase-related protein 2. Biochim Biophys Acta Mol Cell Biol Lipids 1801: 508–516. [Google Scholar]
- Amara S, Delorme V, Record M, et al. 2012. Inhibition of phospholipase A1, lipase and galactolipase activities of pancreatic lipase-related protein 2 by methyl arachidonyl fluorophosphonate (MAFP). Biochim Biophys Acta Mol Cell Biol Lipids 1821: 1379–1385. [Google Scholar]
- Antoniassi R, Miranda PC, Ferreira GF, et al. 2022. Nutritional evaluation of Syagrus coronata kernels and development of cookies prepared with cassava flour and licuri kernels. Food Sci Technol (Campinas) 42: e69720. [Google Scholar]
- Áquila R, Silva J, Silva JV, et al. 2018. The first complete chloroplast genome sequence of Syagrus coronata: comparative analysis in Arecoideae (Arecaceae). Rev Bras Bot 41: 393–401. [Google Scholar]
- Aquino AS, Silva MF, Almeida TS, et al. 2022. Mapping of alternative oilseeds from the Brazilian Caatinga and assessment of catalytic pathways toward biofuels production. Energies 15: 6531. [Google Scholar]
- Araújo PHM, Maia AS, Cordeiro AMTM, et al. 2019. Catalytic deoxygenation of the oil and biodiesel of licuri (Syagrus coronata) to obtain n-alkanes with chains in the range of biojet fuels. ACS Omega 4: 15849–15855. [Google Scholar]
- Aroucha EPTL, Aroucha ML. 2013. Boas práticas de manejo para o extrativismo sustentável do licuri. Brasília: Instituto Sociedade, População e Natureza. [Google Scholar]
- Avelar MHM, Cassimiro DMJ, Santos KC, et al. 2013. Hydrolysis of vegetable oils catalyzed by lipase extract powder from dormant castor bean seeds. Ind Crops Prod 44: 452–458. [Google Scholar]
- Bagaldo AR, Miranda GS, Soares Júnior MS, et al. 2019. Effect of licuri cake supplementation on performance, digestibility, ingestive behavior, carcass traits and meat quality of grazing lambs. Small Rumin Res 177: 18–24. [Google Scholar]
- Barbosa MS, Freire CCC, Almeida LC, et al. 2019. Optimization of the enzymatic hydrolysis of Moringa oleifera Lam oil using molecular docking analysis for fatty acid specificity. Biotechnol Appl Biochem 66: 823–832. [Google Scholar]
- Caro Y, Villeneuve P, Pina M, et al. 2000. Lipase activity and fatty acid typoselectivities of plant extracts in hydrolysis and interesterification. JAOCS J Am Oil Chem Soc 77: 349–354. [Google Scholar]
- Carrière F. 2016. Impact of gastrointestinal lipolysis on oral lipid-based formulations and bioavailability of lipophilic drugs. Biochimie 125: 297–305. [Google Scholar]
- Cavalcanti EDC, Maciel FM, Villeneuve P, et al. 2007. Acetone powder from dormant seeds of Ricinus communis L: lipase activity and presence of toxic and allergenic compounds. Appl Biochem Biotechnol 137–140: 57–65. [Google Scholar]
- Chahinian H, Nini L, Boitard E, Dubès JP, Comeau LC, Sarda L. 2002. Distinction between esterases and lipases: a kinetic study with vinyl esters and TAG. Lipids 37: 653–662. [Google Scholar]
- Chahinian H, Snabe T, Attias C, Fojan P, Petersen SB, Carrière F. 2006. How gastric lipase, an interfacial enzyme with a Ser-His-Asp catalytic triad, acts optimally at acidic pH. Biochemistry 45: 993–1001 [Google Scholar]
- Cibele MASB, Nascimento SR, Alves RC, et al. 2016. Syagrus coronata seed oils have antimicrobial action against multidrug-resistant Staphylococcus aureus. J Med Plants Res 10: 310–317. [Google Scholar]
- Crepaldi IC, Almeida-Muradian LB, Rios MDG, Penteado MDVC, Salatino A. 2001. Composição nutricional do fruto de licuri (Syagrus coronata (Martius) Beccari). Rev Bras Bot 24: 155–159. [Google Scholar]
- da Silva ARC, Soares LRC, Lima AS, Soares CMF, Souza RL. 2022. Strategies to reuse of biocatalysts in the hydrolysis and esterification reactions of licuri (Syagrus coronata (Mart.) Becc.) oil. ChemCatChem 14: 1–10. [Google Scholar]
- da Silva PFF, da Silva RR, Sarubbo LA, Guerra JMC. 2024. Production and optimization of biosurfactant properties using Candida mogii and licuri oil (Syagrus coronata). Foods 13: 1–18. [Google Scholar]
- de Carvalho Alves J, de Souza CO, Santos LM, et al. 2024. Licuri kernel (Syagrus coronata (Martius) Beccari): a promising matrix for the development of fermented plant-based kefir beverages. Foods 13: 2056. [Google Scholar]
- De La Salles KTS, Meneghetti SMP, Ferreira de La Salles W, et al. 2010. Characterization of Syagrus coronata (Mart.) Becc. oil and properties of methyl esters for use as biodiesel. Ind Crops Prod 32: 518–521. [Google Scholar]
- de Oliveira Alves JV, Barbosa BVD, Aguiar IFS, et al. 2024. Fixed oil from the Caatinga palm Syagrus coronata with effect on complete Freund’s adjuvant (CFA)-induced arthritis. Inflammopharmacology 32: 3389–3398. [Google Scholar]
- de Souza dos Santos GE, Ide AH, Duarte JLS, McKay G, Sousa Silva AO, Meili L. 2020. Adsorption of anti-inflammatory drug diclofenac by MgAl/layered double hydroxide supported on Syagrus coronata biochar. Powder Technol 364: 229–240. [Google Scholar]
- Delorme V, Dhouib R, Canaan S, Fotiadu F, Carrière F, Cavalier JF. 2011. Effects of surfactants on lipase structure, activity, and inhibition. Pharm Res 28: 1831–42. [Google Scholar]
- Dhouib R, Laroche-Traineau J, Shaha R, et al. 2011. Identification of a putative triacylglycerol lipase from papaya latex by functional proteomics. FEBS J 278: 97–110. [Google Scholar]
- dos Santos Souza TG, da Silva MM, Feitoza GS, et al. 2021. Biological safety of Syagrus coronata (Mart.) Becc. fixed oil: cytotoxicity, acute oral toxicity, and genotoxicity studies. J Ethnopharmacol 272: 113941. [Google Scholar]
- Ejedegba BO, Onyeneke EC, Oviasogie PO. 2007. Characteristics of lipase isolated from coconut (Cocos nucifera Linn) seed under different nutrient treatments. Afr J Biotechnol 6: 723–727. [Google Scholar]
- Farzaneh V, Bakhshabadi H, Gharekhani M, et al. 2017. Application of an adaptive neuro-fuzzy inference system (ANFIS) in the modeling of rapeseeds' oil extraction. J Food Process Eng 40: 1–8. [Google Scholar]
- Ferrato et al., 1997, cited on page 11, line 513 is missing in the reference list. Here is the complete reference to be added: Ferrato F, Carrière F, Sarda L and Verger R. (1997) A critical re-evaluation of the phenomenon of “Interfacial Activation”. Methods in Enzymology 286: 327–347. [Google Scholar]
- Francisconi AF, Morales Marroquín JA, Cauz-Santos LA, et al. 2023. Complete chloroplast genomes of six neotropical palm species, structural comparison, and evolutionary dynamic patterns. Sci Rep 13: 1–15. [CrossRef] [PubMed] [Google Scholar]
- Huerta-Yépez S, Tirado-Rodriguez AB, Hankinson O. 2016. Role of diets rich in omega-3 and omega-6 in the development of cancer. Bol Med Hosp Infant Mex 73: 446–456. [Google Scholar]
- Iha OK, Alves FCSC, Suarez PAZ, et al. 2014. Physicochemical properties of Syagrus coronata and Acrocomia aculeata oils for biofuel production. Ind Crops Prod 62: 318–322. [Google Scholar]
- Jha PK, Kudachikar VB, Kumar S. 2013. Lipase inactivation in wheat germ by gamma irradiation. Radiat Phys Chem 86: 136–139. [Google Scholar]
- Kouteu PAN, Blin J, Baréa B, Barouh N, Villeneuve P. 2017. Solvent-free biodiesel production catalyzed by crude lipase powder from seeds: effects of alcohol polarity, glycerol, and thermodynamic water activity. J Agric Food Chem 65: 8683–8690. [Google Scholar]
- Leal LB, Sousa GD, Seixas KB, De Souza PHN, De Santana DP. 2013. Determination of the critical hydrophile-lipophile balance of licuri oil from Syagrus coronata: application for topical emulsions and evaluation of its hydrating function. Braz J Pharm Sci 49: 167–173. [Google Scholar]
- Lisboa MC, Wiltshire FMS, Fricks AT, et al. 2020. Oleochemistry potential from Brazil northeastern exotic plants. Biochimie 178: 96–104. [Google Scholar]
- Mattos C, Andrade J, Oliveira PCO, Veloso MCC, Romeiro GA, Chagas EF. 2021. Fumigant activity of pyrolysis fractions derived from waste licuri's pericarp (Syagrus coronata - Arecaceae) in the control of the red flour beetle Tribolium castaneum (Herbst). Environ Chall 5: 100294. [Google Scholar]
- Morcillo F, Cros D, Billotte N, et al. 2013. Improving palm oil quality through identification and mapping of the lipase gene causing oil deterioration. Nat Commun 4: 2160. [Google Scholar]
- Mustafa A, Faisal S, Ahmed IA, et al. 2023. Has the time finally come for green oleochemicals and biodiesel production using large-scale enzyme technologies? Current status and new developments. Biotechnol Adv 69: 108275. [Google Scholar]
- Ngando Ebongue GF, Dhouib R, Carrière F, et al. 2006. Assaying lipase activity from oil palm fruit (Elaeis guineenses Jacq.) mesocarp. Plant Physiol Biochem 44: 611–617. [Google Scholar]
- Ngando Ebongue GF, Koona P, Nouy B, et al. 2008. Identification of oil palm breeding lines producing oils with low acid values. Eur J Lipid Sci Technol 110: 505–509. [Google Scholar]
- Oliveira VB, Yamada LT, Fagg CW, et al. 2012. Native foods from Brazilian biodiversity as a source of bioactive compounds. Food Res Int 48: 170–179 [Google Scholar]
- Pereira RAG, Oliveira CJB, Medeiros AN, et al. 2010. Physicochemical and sensory characteristics of milk from goats supplemented with castor or licuri oil. J Dairy Sci 93: 456–462. [Google Scholar]
- Pierozan MK, Da Costa RJ, Antunes OAC, et al. 2009. Optimization of extraction of lipase from wheat seeds (Triticum aestivum) by response surface methodology. J Agric Food Chem 57: 9716–9721. [Google Scholar]
- Ramos DG, Barbosa Cardoso KB, Liu TPSL, et al. 2025. Biotechnological potential of licuri (Syagrus coronata) cake for sustainable production of multienzyme extract via submerged fermentation. Waste Biomass Valorization: in press. [Google Scholar]
- Ramos DG, Lima JMS, Barbosa Filho JPM, et al. 2025. Licuri (Syagrus coronata) oil cake as a substrate for collagenolytic protease production by submerged fermentation using a novel strain of Penicillium citrinum. Waste Biomass Valorization 16: 1261–1276. [Google Scholar]
- Rivera I, Robles M, Mateos-Díaz JC, et al. 2017. Functional expression, extracellular production, purification, structure modeling and biochemical characterization of Carica papaya lipase 1. Process Biochem 56: 109–116. [Google Scholar]
- Rodrigues CA, Barbosa MS, Dos Santos JCB, et al. 2021. Computational and experimental analysis on the preferential selectivity of lipases for triglycerides in licuri oil. Bioprocess Biosyst Eng 44: 2141–2151. [Google Scholar]
- Sagiroglu A, Arabaci N. 2005. Sunflower seed lipase: extraction, purification, and characterization. Prep Biochem Biotechnol 35: 37–51. [Google Scholar]
- Sahaka M, Amara S, Lecomte J, et al. 2021. Quantitative monitoring of galactolipid hydrolysis by pancreatic lipase-related protein 2 using thin layer chromatography and thymol-sulfuric acid derivatization. J Chromatogr B 1173: 122674. [Google Scholar]
- Sanchez S, Demain AL. 2011. Enzymes and bioconversions of industrial, pharmaceutical, and biotechnological significance. Org Process Res Dev 15: 224–230 [Google Scholar]
- Santos KC, Cassimiro DMJ, Avelar MHM, et al. 2013. Characterization of the catalytic properties of lipases from plant seeds for the production of concentrated fatty acids from different vegetable oils. Ind Crops Prod 49: 462–470. [Google Scholar]
- Santos LMM, Nascimento JS, Santos MAG, et al. 2017. Fatty acid-rich volatile oil from Syagrus coronata seeds has larvicidal and oviposition-deterrent activities against Aedes aegypti. Physiol Mol Plant Pathol 100: 35–40. [Google Scholar]
- Santos LER, Meili L, Soletti JI, et al. 2020. Impact of temperature on vacuum pyrolysis of Syagrus coronata for biochar production. J Mater Cycles Waste Manag 22: 878–886 [Google Scholar]
- Scalet V, Da Róz AL, Santos LRO, et al. 2019. Waste of the licuri (Syagrus coronata) nut shells: an alternative energy source. Rev Bras Energ Renováveis 8: 583–597. [Google Scholar]
- Sehgal S, Sharma V. 2020. Palm/palm kernel (Elaeis guineensis). In: Oilseeds: Health Attributes and Food Applications. Singapore: Springer Singapore. [Google Scholar]
- Silva SI, Oliveira AFM, Negri G, et al. 2014. Seed oils of Euphorbiaceae from the Caatinga, a Brazilian tropical dry forest. Biomass Bioenergy 69: 124–134. [Google Scholar]
- Sommo M, Andrade de Aguiar L, Raposo A, et al. 2024. Development and rapid sensory descriptive characterization of cereal bars made with Brazilian licuri nut (Syagrus coronata). Foods 13: 1–13. [Google Scholar]
- Sorigué D, Légeret B, Cuiné S, et al. 2017. An algal photoenzyme converts fatty acids to hydrocarbons. Science 357: 903–907 [Google Scholar]
- Souza Dos Santos B, Bezerra Filho CM, Nascimento Junior, et al. 2019. Anti-staphylococcal activity of Syagrus coronata essential oil: biofilm eradication and in vivo action on Galleria mellonella. Microb Pathog 131: 150–157. [Google Scholar]
- Tieves F, Hollmann F. 2025. Enzymatic valorization of fatty acids in oleochemical synthesis. Eur J Lipid Sci Technol 127: e70000. [Google Scholar]
- Tiss A, Carrière F, Verger R. 2001. Effects of gum Arabic on lipase interfacial binding and activity. Anal Biochem 294: 36–43 [Google Scholar]
- USP. 2018. ‘<1025> Pancreatin.’ in Inc. The United States Pharmacopeial Convention (ed.), The United States Pharmacopeia and National Formulary USP 41-NF 36 (The United States Pharmacopeial Convention, Inc.: Rockville, MD). [Google Scholar]
- Victor MM, Uchoa JE, Riatto VB. 2017. First use of cactus and cushcush as biocatalysts in the enantioselective reduction of β-keto esters. Ind Crops Prod 96: 126–131. [Google Scholar]
- Villeneuve P. 2003. Plant lipases and their applications in oils and fats modification. Eur J Lipid Sci Technol 105: 308–317. [Google Scholar]
- Wancura JHC, Cipolatti EP, Manoel EA, et al. 2025. Advanced biocatalytic processes for the conversion of renewable feedstocks into high-value oleochemicals. Catalysts 15: 600. [Google Scholar]
- Wattanakul J, Sahaka M, Amara S, et al. 2019. In vitro digestion of galactolipids from chloroplast-rich fraction of plant residues and spinach leaves. Food Funct 10: 7806–7817. [CrossRef] [PubMed] [Google Scholar]
- Xavier G, De Paula Filho, Barreira TF, et al. 2015. Study of the physical and physicochemical characteristics of fruits of the licuri palm (Syagrus coronata) found in the Atlantic Forest of Minas Gerais, Brazil. Food Sci Technol 35: 474–480. [Google Scholar]
- Zhang Y, Launay H, Schramm A, et al. 2018. Exploring intrinsically disordered proteins in Chlamydomonas reinhardtii. Sci Rep 8: 6805. [Google Scholar]
Cite this article as: Milena Chagas Lisboa, César De Almeida Rodrigues, Moulay Sahaka, Hélène Launay, Ranyere Lucena de Souza, Frederic Carriere, Alvaro Silva Lima, Cleide Mara Faria Soares. 2026. Fruit mesocarp of the Brazil-native Licuri palm (Syagrus coronata) as a dual and sustainable source of oil and lipase for biorefinery applications. OCL 33: 20. https://doi.org/10.1051/ocl/2026011
All Tables
Lipid extraction yields (% w/w) from Licuri kernel and mesocarp using hexane in a Soxhlet apparatus (with prior drying in oven or desiccator) or cold hexane. Values are mean ± SD (n = 3) and are expressed on a dry basis.
Comparison of fatty acid composition (relative% w/w) of kernel and mesocarp of licuri and oil palm fruits. afrom (Iha et al., 2014); bfrom (Sehgal and Sharma, 2021).
Lipase activities (U·/g−) in the powders obtained from kernel (DDK) and mesocarp (DDM) from licuri fruit. Activities were measured at pH 7 using either olive oil (3% gum Arabic) or tributyrin as substrates.
Protein matches with high similarity to lipolytic enzymes obtained from the peptide mass fingerprint of PPLL protein bands separated by SDS-PAGE. Species names: Cocos Nucífera, coconut; Elaeis guineensis, oil palm; Carica papaya, papaya; Phoenix dactylifera, date palm. PSM, number of peptide single matches.
All Figures
|
Fig 1 Geographical distribution of the licuri palm tree (Syagrus coronata (Mart.) Becc.) and anatomy of its fruit. Licuri is endemic to regions from northern Minas Gerais to southern Pernambuco (panel A). Panel B shows the standard shape and color of the licuri fruit used in this study, along with anatomical features, cross-section, and physical characteristics of ripe, freeze-dried licuri fruits collected from Tingui Farm in Malhada dos Bois, Sergipe, Brazil (Latitude: 10°20′58″ S; Longitude: 36°55′27″ W). |
| In the text | |
|
Fig 2 Diagram of the preparation process for the preliminary characterization of licuri lipase. (A) Preparation of dry delipidated mesocarp (DDM). (B) Preparation of pre-purified licuri lipase (PPLL), including freeze-drying of pooled liquid and gelatinous phases. |
| In the text | |
|
Fig 3 Thin-layer chromatography analysis of lipids extracted from the Licuri mesocarp (Mes) and endocarp (End) with cold hexane. (A) Separation of neutral lipids stained with copper acetate-phosphoric acid. DAG, Diacylglycerol (Rf: 1.3-DAG: 0.31 ± 0.02; 1.2(2.3)-DAG: 0.25 ± 0.02); End, endocarp or kernel; FFA, Free fatty acids (Rf: 0.48 ± 0.03); MAG, Monoacylglycerol (Rf: 0.02 ± 0.01); Mes, Mesocarp; Rs, Reference standards including triolein, oleic acid, diolein, and monoolein; TAG, Triacylglycerol (Rf: 0.88 ± 0.05). (B) Separation of polar lipids stained with thymol-sulfuric acid. End, endocarp; MGDG: monogalactosyl diacylglycerol (Rf: 0.63 ± 0.05); DGDG: digalactosyl diacylglycerol (Rf: 0.31 ± 0.02). (C) Hydrolysis of MGDG and DGDG reference standards by pancreatic lipase-related protein 2 (PLRP2), leading to the formation of monogalactosyl monoacylglycerol (MGMG; Rf: 0.40 ± 0.03) and digalactosyl monoacylglycerol (DGMG; Rf: 0.10 ± 0.02). The samples were obtained according to Sahaka et al. (2021). Original pictures are available in the Supporting Information (Figs. S1, S2 and S3). |
| In the text | |
|
Fig 4 Thin-layer chromatography analysis of lipid extracts from licuri mesocarp obtained with and without blanching. (A) Polar lipids stained with thymol-sulfuric acid. (Rf: FFA: 0.69 ± 0.05; MGDG: 0.63 ± 0.05; MGMG: 0.40 ± 0.03; DGDG: 0.31 ± 0.02; DGMG: 0.10 ± 0.02). (B) Non-polar lipids stained with copper acetate-phosphoric acid. FFA (Rf: 0.62 ± 0.05); MGDG: monogalactosyl diacylglycerol (Rf: 0.54 ± 0.05); DGDG: digalactosyl diacylglycerol (Rf: 0.29± 0.02). Original pictures are available at Supporting information (Figs. S4 and S5). |
| In the text | |
|
Fig 5 Stability of licuri mesocarp lipase. (A) Variation with time in the lipase activity of licuri mesocarp homogenates and dry delipidated mesocarp (DDM) dispersion kept at 4°C. (B) Residual activity of PPLL after incubation at either room temperature (with and without protease inhibitors) or 4°C. Lipase activity was measured using tributyrin (TC4) as a substrate at pH 7. |
| In the text | |
|
Fig 6 Effect of several parameters on lipase activity in licuri mesocarp. (A) Variation with pH in the lipase activity on olive oil in the presence of 3% w/v gum arabic. (B) Effect of Gum Arabic (GA) concentration on the hydrolysis of olive oil at pH 8. (C) Effect of bile salts (NaTDC) concentration on tributyrin hydrolysis reaction at pH 7. |
| In the text | |
|
Fig 7 Specific activity of pre-purified licuri lipase extract (PPLL) on various lipid substrates. (A) pH-dependent activities on olive oil, licuri oils tributyrin, phospholipids (Egg PC), and vinyl ester. Vinyl propionate was used at a concentration below its solubility in water (<86 mM; 100 μL/30 mL per assay) to measure the esterase activity of licuri lipase on a soluble substrate. (B) Interfacial activation: PPLL activity on vinyl propionate at substrate concentrations below (30 mM) and above (153 mM) the saturation (86 mM). |
| In the text | |
|
Fig 8 SDS–PAGE analysis of PPLL. Black arrows indicate the various protein bands present in LLPP. The red arrow indicates the material remaining on the stacking gel. |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.