")
")
| Issue |
OCL
Volume 32, 2025
Contaminants in oils and fats / Contaminants des huiles et corps gras
|
|
|---|---|---|
| Article Number | 22 | |
| Number of page(s) | 13 | |
| DOI | https://doi.org/10.1051/ocl/2025019 | |
| Published online | 16 July 2025 | |
Review
Cadmium and other metallic contaminants in cacao: Update on current knowledge and mitigation strategies☆
Cadmium et autres contaminants métalliques dans le cacao : Mise à jour des connaissances actuelles et des stratégies d'atténuation
1
Université Grenoble Alpes, Université Savoie Mont Blanc, CNRS, IRD, Université G. Eiffel, ISTerre, Grenoble, France
2
ESRF, The European Synchrotron, Grenoble, France
3
Escuela Superior Politécnica del Litoral, ESPOL, Facultad de Ciencias de la Vida, Campus Gustavo Galindo Km. 30.5 Vía Perimetral, P.O. Box 09-01-5863, Guayaquil, Ecuador
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
Received:
7
March
2025
Accepted:
17
June
2025
Abstract
Cadmium (Cd) is a non-essential metal classified as class 1 carcinogen, which can induce detrimental effects for humans. Food is the main route of Cd exposure for the general population (except smokers). Chocolate products account for 4% and 9% of the dietary Cd intake for adults and children, respectively, in Europe. Since 2019, the EU limits Cd concentration in chocolate. The maximum permitted concentration (MPC) varies from 0.30 to 0.80 mg Cd kg−1 depending on the% of total cocoa solids in the final product. Cacao (Theobroma cacao L.) producers in Latin America and the Caribbean, who provide 19.4% of global cacao production, are strongly impacted because a significant fraction of their production exceeds these limits. In this review, the current knowledge on the factors influencing the soil Cd enrichments and Cd phytoavailability is discussed. The Cd transfer to cacao trees and to beans, and the responses of cacao trees to Cd exposure are also presented. Then, various mitigation strategies, including post-harvest treatments, soil amendments, microbial remediation (or bioremediation), breeding programs and grafting, are summarized. We also discuss the transfer of knowledge to stakeholders in the cacao supply chain. Finally, current knowledge on the presence of other metallic contaminants in cacao beans, particularly nickel (Ni), is reviewed.
Résumé
Le cadmium (Cd) est un métal non essentiel classé comme cancérogène de classe 1, qui peut avoir des effets néfastes sur l’Homme. L'alimentation est la principale voie d'exposition au Cd pour la population générale (à l’exception des fumeurs). En Europe, les produits à base de chocolat représentent 4% et 9% de l'apport alimentaire en Cd chez les adultes et les enfants, respectivement. Depuis 2019, l'UE limite la concentration de Cd dans le chocolat. La limite maximale pour cette concentration varie de 0,30 à 0,80 mg Cd kg−1 selon le pourcentage de cacao solide total dans le produit final. Les producteurs de cacao (Theobroma cacao L.) d'Amérique latine et des Caraïbes, qui fournissent 19,4% de la production mondiale de cacao, sont fortement impactés puisqu'une fraction importante de leur production dépasse ces limites. Dans cette revue, les connaissances actuelles sur les facteurs influençant l'enrichissement en Cd du sol et la phytodisponibilité du Cd sont discutées. Le transfert du Cd vers les cacaoyers et les fèves, ainsi que les réponses des cacaoyers à l’exposition au Cd sont aussi présentés. Les stratégies d'atténuation, en particulier les traitements après récolte, l’apport d’amendements au sol, la remédiation microbienne ou bioremédiation, la sélection de variétés et le greffage, sont ensuite résumés. Nous discutons également du transfert de connaissances aux parties prenantes de la chaîne d'approvisionnement en cacao. Enfin, les connaissances actuelles sur la présence d'autres contaminants métalliques dans les fèves de cacao, en particulier le nickel (Ni), sont présentées.
Key words: Theobroma cacao L. / Cocoa / cadmium / mitigation / metallic contaminant
Mots clés : Theobroma cacao L. / Cacao / cadmium / atténuation / contaminant métallique
Contribution to the Topical Issue: “Contaminants in oils and fats / Contaminants des huiles et corps gras”.
© F. Letort et al., Published by EDP Sciences, 2025
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Highlights
The Cd source in cacao from Latin America is mostly geogenic and arises from sedimentary rocks.
Soil amendment-based mitigation was so far unsuccessful to reduce bean Cd, and most promising strategies include post-harvest treatments and cultivar breeding.
Genetic-based mitigation requires a screening on Cd accumulation phenotypes, and a knowledge of the ecophysiology of Cd in cacao.
Adaptation strategies should consider Cd together with environmental stress parameters for cacao.
1 Introduction
The global annual cacao production in 2022 was estimated at 5.6 million tons (FAOSTAT, 2022). Africa is the highest producer with 67.9%, followed by Latin America and the Caribbean (19.4%) and Asia and Oceania (12.5%) (FAOSTAT, 2022). Ivory Coast is the largest exporter of cacao beans (42% of the world production), followed by Ghana (12%), Indonesia (12%) and Ecuador[CE1] (6%) (Fig. 1).
Cadmium is a non-essential metal classified as class 1 carcinogen (WHO/IARC, 2025). Exposure of the human population at low levels can result in chronic toxicity. Food is the main route of Cd exposure for the general population, except for smokers. The main contributors of Cd in the human diet are cereals, potatoes and seafood, but chocolate products account for 4% and 9% of the dietary Cd intake for adults and children, respectively (EFSA, 2012). Therefore, the EU set limits on Cd concentration in chocolate products in 2019, to complement the previous limits on other foodstuffs. These limits are 0.10 mg Cd kg−1 for chocolates with less than 30% dry cacao solids, 0.30 mg Cd kg−1 for chocolates with 30% to 50% dry cacao solids, 0.80 mg Cd kg−1 for chocolates with more than 50% dry cacao solids, and 0.60 mg Cd kg−1 for cacao powder (EC, 2014). The Codex Alimentarius Commission (CAC) has adopted the same limits (Codex Alimentarius Commission, 2018). The cacao processing industry has translated the EU limits into maximum Cd levels in fermented beans between 0.50 and 1.10 mg Cd kg−1 (Vanderschueren et al., 2021).
Cacao beans from Latin America have higher Cd concentrations than those from Africa or Asia (Meter et al., 2019). Therefore, the EU regulation on Cd in chocolate has socio economic impacts in cacao exporting countries such as Ecuador. A study estimated the total economic cost of abandoning cacao cultivation in areas of Ecuador with bean Cd above 0.60 mg Cd kg−1 at about 750 million US$ (Vázquez-deCastro et al., 2024). In countries with local consumption of the cacao production, it is also important to assess and control the Cd level in beans.
This Cd issue has triggered intense research to assess the origin of the contamination and to propose mitigation strategies. Two authors of this review co-authored a review in 2021 (Vanderschueren et al., 2021). This review proposes a short update, mostly based on studies published since 2021.
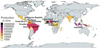 |
Fig. 1 Production of cacao beans per country in tons in 2022. Only countries ≥ 800 tons were taken into consideration (FAOSTAT, 2022). |
2 Origin and extent of the cadmium occurrence in Latin America
2.1 Origin(s) of soil cadmium occurrence
Understanding the origins of soil Cd and bean Cd is a prerequisite to design mitigation strategies. High soil Cd can have a natural and an anthropogenic origin. Natural (geogenic) Cd enrichment in soils generally results from high geological background and Cd incorporation during pedogenic processes. Anthropogenic soil Cd contaminations generally result from mining, smelting and other industrial activities, traffic, and application of phosphate (P) fertilizers, biosolids and irrigation with wastewaters (McLaughlin et al., 2021). A geogenic enrichment of the soils is likely the main explanatory factor for the high total soil Cd in cacao fields in Latin America for several reasons: (1) Cadmium maps from national surveys (see next paragraph) show a relationship between Cd and soil type and geology, with Cd enrichments in soils developed on sedimentary bedrock (2) High soil Cd impact the cacao production in Latin America, but not in Africa (although agricultural practices do not strongly differ) (3) The majority of cacao is produced in small-scale farms (Meter et al., 2019; Vanderschueren et al., 2021) (generally < 5 ha) with low supplies, thus reducing the risk of Cd input via commercial P fertilizers. However, the use of low-cost amendments such as biosolids cannot be excluded.
The soil Cd enrichment in Latin America was suggested to result from the deposition of alluvial sediments enriched in Cd (Meter et al., 2019; Maddela et al., 2020; Vanderschueren et al., 2021). A recent study identified parent rocks of sedimentary origin, such as limestone, marl and black shale, as a source of Cd in cacao field soils of Santander, Colombia (Joya-Barrero et al., 2023). In another study, hydrothermally altered rocks were suggested as a source of Cd in cacao field soils of Piura, North Peru (Guarín et al., 2023). Cadmium concentration generally increases in the order subsoil < topsoil (Vanderschueren et al., 2021; Blommaert et al., 2025) and subsoil < topsoil < litter (Gil et al., 2022; Guarín et al., 2024). Thus, the soil Cd enrichment probably originates from the weathering of the underlying bedrock and from alluvial deposits, and the topsoil is progressively enriched due to Cd accumulation in cacao leaves and their incorporation after leaf fall (Gramlich et al., 2018). In addition to geogenic enrichment, local hotspots may result from anthropogenic activities (Argüello et al., 2019; Meter et al., 2019; Maddela et al., 2020; Vanderschueren et al., 2021). Soil and bean Cd contamination due to mining activities were also reported in Africa (Ghana), with soil containing up to 6.31 mg Cd kg−1, and beans containing up to 3.16 mg Cd kg−1 (Frimpong et al., 2025). The use of mineral P-fertilizer could be an additional source of Cd. Variable Cd concentrations were reported in fertilizers available in Latin America, from 3 to 30 mg Cd kg−1 P205 in Colombia (Bravo et al., 2022) and from 30 to 120 mg Cd kg−1 P205 in Honduras and Peru (Gramlich et al., 2017; Guarín et al., 2023). As a comparison, the current Cd limit for P fertilizers in the EU is 60 mg Cd kg−1 P205. Although some authors suggest that P fertilizers may be a significant source of Cd for cacao plants (Barraza et al., 2017; Guarín et al., 2023), other authors consider that this source is marginal because the use of fertilizers in cacao plantations in Latin America is limited (Vanderschueren et al., 2021).
2.2 Nation-scale surveys in Latin America
Nationwide surveys on soils and beans Cd were carried out in several countries in Latin America (Tab. 1). They showed a large variability in bean Cd, especially in Colombia and Ecuador (Standard deviation (SD): 2.71 and 1.16 mg Cd kg−1 larger than the mean: 2.17, and 0.9 mg Cd kg−1, respectively). This variability results mainly from regional differences. Regions with highest soil Cd correspond to areas with calcareous soils, in which the weathering of Cd-rich rocks releases Cd-rich alluvial deposits, which can be deposited on soils (Bravo et al., 2024). In addition to this variability between geological subunits (e.g., Gramlich et al., 2018), a variability within a single field was observed (Vanderschueren et al., 2021). Finally, strong local heterogeneities within the same cacao plantation were observed (e.g., Letort et al., 2025).
3 Factors affecting Cd concentration in beans
3.1 Soil parameters, microorganisms and cropping system
Based on a meta-analysis of 780 cacao soil-plant samples from field surveys, total soil Cd, pH, and soil organic carbon (SOC) accounted for 41% of bean Cd variability. Bean Cd increased proportionally with total soil Cd, rose 1.5 times per unit decrease in pH, and was reduced 1.8 times when SOC doubled (Vanderschueren et al., 2021). Low soil pH increases Cd solubility and thus its availability, by enhancing competition between H+ and Cd2+ for organic matter, clay, Fe and Mn surfaces binding sites. Conversely, at high soil pH, increased negative charges in the soil promote Cd sorption, reducing its uptake by plants (Smolders and Mertens, 2013). Wade et al. (2022) performed another meta-analysis based on 31 studies containing soil Cd parameters (pH, SOC, total and phytoavailable Cd) and plant Cd (leaf Cd and bean Cd). Total soil Cd and pH, along with leaf Cd, were the best predictors of bean Cd concentration. Total soil Cd was identified as the primary driver of bean Cd, while higher pH and SOM (Soil Organic Matter) reduced Cd bioaccumulation in beans (Vanderschueren et al., 2021). By contrast, based on a survey of 150 farms in Costa Rica, Carrillo et al. (2023) found that total soil Cd alone explained ∼58% of bean Cd variability, total soil Cd and SOC explained 62%. Soil pH had no statistical effect. A plausible explanation is the relative homogeneity of soil pH among these 150 farms in Costa Rica (5.53 ± 0.60). A field study of 30 cacao farms in northern Colombia (Guarín et al., 2024) revealed significant positive correlations between total soil, leaf, and litter Cd with total soil Zn (R² = 0.50, 0.51, and 0.55, respectively), consistent with (Arévalo-Gardini et al. (2017). However, Vanderschueren et al. (2021) did not identify total soil Zn as a significant factor to predict bean Cd. Guarín et al. (2024) also found no significant correlation between soil Cd and pH and only a weak relationship with SOM, likely due to low Cd levels and variability. Further studies on soil parameters, such as salinity and micronutrients (Fe, Mn, and Zn) availability are required to clarify their interaction with Cd and their influence on its accumulation in cacao.
Phytoavailability of Cd is also modulated by biological factors such as microorganisms. Rhizosphere bacteria and fungi are pivotal in the fluxes of nutrients, water and xenobiotics in the plant. However, their impact on Cd availability and uptake in cacao crops in field conditions remain unclear. Cáceres et al. (2021) characterized bacterial communities in Colombian cacao farms and identified a number of Cd-tolerant strains mostly belonging to the Proteobacteria phylum. The strains identified included Burkholderia sp., Enterobacter sp., Escherichia sp., Klebsiella sp., Ralstonia sp., Serratia sp., Exiguobacterium sp., Dermacoccus sp., Lactococcus sp., and Staphylococcus sp., some of them being commonly found in metal-contaminated soils. Bacterial community composition was significantly correlated with soil pH, SOC and total Cd. In another study on rhizosphere soil samples from Colombian cacao farms, a link between the bacterial community composition and DTPA-extractable Cd was evidenced (Jaramillo-Mazo et al., 2024). Further studies are needed to verify whether this link is causal. Twenty-six Cd-tolerant strains belonging to 14 genus, were isolated from cacao farm soil in Colombia containing 1.16 to 3.74 mg Cd kg−1 when exposed to 6, 12 and 24 mg L−1 of CdCl2 in petri dish (Bravo et al., 2018). On the other hand, a study conducted at the International Cocoa Genebank, Trinidad (ICGT) identified four prokaryotic and fungal taxa whose relative abundance was positively correlated with plant Cd (Schmidt et al., 2025). Further studies should be conducted to evaluate whether this correlation implies a causal link. Further research would be necessary to unveil the interactions between soil microorganisms and soil Cd of cacao plantations, and the net impact on Cd availability, as a function of soil properties and environmental conditions.
The effect of drought stress on Cd accumulation in six-month-old cacao plantlets was evaluated in a pot experiment on three genotypes (IMC-67 x PA-121, SCA-6 x PA-121 and IMC-67) grown on soil containing 0.356 mg Cd kg−1, after 19 and 27 days (Ortiz-Álvarez et al., 2023). Water deficit increased Cd concentration in the whole plant in IMC-67 x PA-121 on day 19 and 27 and in SCA-6 x PA-121 on day 27. In addition, water deficit increased Cd translocation from roots to leaves in IMC-67 x PA-121 and IMC-67, while there were no significant changes in SCA-6 x PA-121.
Finally, the influence of agronomic practices (monoculture vs. agroforestry) on cacao Cd was assessed. Argüello et al. (2019) reported no significant effect of the use of mineral N-P-K fertilizers between these two practices. Gramlich et al. (2017) found a higher leaf Cd in monoculture compared to agroforestry. They suggested that higher plant density could cause higher competition for nutrients, decreasing cacao growth and capacity to accumulate Cd. Vallejos-Torres et al. (2023) evaluated the influence of culture management on Cd uptake and mycorrhization, in Peru. Enhancement in both foliar and beans Cd in monoculture compared to agroforestry was determined, in addition to decreased length of extraradical mycelium. The authors hypothesized that plant biodiversity induced a better root system, which resulted in improved habitat for arbuscular mycorrhizal fungi and therefore reduced Cd uptake in cacao plants. In conclusion, the effectiveness of agroforestry in reducing bean Cd remains uncertain, requiring further research.
3.2 Genetics
Genetic variations in the bioaccumulation of Cd in cacao trees was first demonstrated by Lewis et al. (2018), based on 100 native cultivars belonging to 10 genetic groups, growing at ICGT (International Cocoa Genebank, Trinidad), on soil with similar phytoavailable Cd. A 13-fold variation in bean Cd and a 7-fold variation in leaf Cd between accessions was observed. This genetic variability in Cd accumulation was confirmed in other studies. Blommaert et al. (2025) compared high and low Cd-accumulating cacao cultivars growing in the same ICGT plot as in Lewis et al. (2018). The high Cd accumulator exhibited 2.9-fold higher total Cd plant uptake than the low Cd accumulator. Interestingly, the distribution among organs was similar, suggesting that cultivar differences are at the root uptake stage, not at the translocation stage. Still in the same ICGT plot, Schmidt et al. (2025) measured leaf and stem Cd in 16 genotypes classified as high and low Cd accumulators by Lewis et al. (2018). They reported a wide range of leaf Cd, with a factor ∼30 between the two extremes. The authors suggested that this continuous distribution could indicate the involvement of multiple genes in Cd uptake and translocation. Arévalo-Hernández et al. (2021) evaluated genetic variation among 53 wild and domesticated cacao genotypes from different Latin American countries grown on Cd-spiked soil with 25 mg Cd kg−1 (not an environmentally relevant concentration). After 4 months in greenhouse conditions, juvenile cacao plants showed distinctive Cd uptake patterns, with two orders of magnitude in shoot Cd between the low and high Cd accumulators. Among genotypes, CCN-51 showed shoot Cd (2 mg kg−1) belonging to the cluster of lowest shoot Cd (1.38 ± 0.22 mg kg−1). Moore et al. (2020) using a hydroponic experiment tested 19 cacao genotypes subjected to 20 μmol Cd L−1 (2.25 mg Cd L−1) (not an environmentally relevant concentration) from International Cocoa Quarantine Centre (ICQC). Total Cd accumulated by cacao plants varied with an extremum factor of ∼9 among genotypes. Plantlets also exhibited isotope fractionation from solution to total plant (Δ114/110 Cdtot-sol) ranging from −0.34 to 0.01‰, which may suggest different Cd uptake transporters.
This genetic variation has been considered for selecting rootstocks that may be used to decrease bean Cd (Borjas-Ventura et al., 2022; Galvis et al., 2023). EET-61, EET-62, IMC-60, IMC-67, PA-121, PA-150, PA-46, SCC-85, and SCC-86 genotypes of juvenile stage from the Germplasm Bank of the La Suiza Research Center were evaluated for rootstock potential during 90 days exposure in a Cd-spiked soil at 0, 6, and 12 mg Cd kg−1 (strongly contaminated soil). Cadmium concentration was measured in leaves, stems, and roots. Results at 6 mg Cd kg−1 showed that EET-61 had the highest plant Cd concentration in plant with a factor ∼11 compared to the lowest, IMC-67 and PA-46. The authors proposed that these latter cultivars could be used as rootstocks for reducing the translocation of Cd from roots to shoots (Galvis et al., 2023). In Peru, Borjas-Ventura et al. (2022) analyzed Cd absorption and its impacts by comparing 6 genotypes differing by morphological and physiological parameters: roots were exposed in a Cd-spiked soil from 50 to 150 mg Cd kg−1 (not an environmentally relevant concentrations). At the highest concentration, POUND-7 absorbed the greatest total amount of Cd mostly accumulated in roots, around three times more than in the lowest genotype CCN-51, which accumulated Cd mainly in leaves. Hence, despite unrealistic exposure conditions, POUND-7 could have a potential for rootstock because of a high retention of Cd in roots. Finally, different plant genotypes may modulate the microbial composition in the rhizosphere (Schmidt et al., 2025), and then potentially affect Cd availability, as discussed in section 3.1.
4 Uptake, translocation and partitioning of cadmium within the cacao tree
Cadmium ion (Cd2+) has similar properties as the main micronutrients Zinc (Zn2+), Iron (Fe2+), and Manganese (Mn2+), and can be transferred in the plant by the transporters of these essential elements (Clemens and Ma, 2016). The transporter TcNRAMP5, belonging to the Natural Resistance Associated Macrophage Proteins (NRAMPs) family, likely plays a role in the root uptake of Cd by cacao (Ullah et al., 2018; Moore et al., 2020). The reader is referred to a previous review for more details (Vanderschueren et al., 2021). There can be a competition between Cd and essential metals for their root uptake. In a pot experiment with a Cd and Zn-spiked soil, increased soil Zn decreased Cd uptake in cacao plants (dos Santos et al., 2020). This result could suggest that other transporters, such as ZIP, could be involved in Cd uptake in cacao. Further studies are necessary to test this hypothesis. In contrast, in a similar experiment with Cd and Mn-spiked soil, increased soil Mn increased Cd uptake and partly alleviated Cd toxicity (Barroso et al., 2023). Besides competition with micronutrients in sufficient or excess conditions potentially leading to reduced plant Cd uptake, micronutrient deficiencies could enhance plant Cd uptake (Sterckeman and Thomine, 2020). This hypothesis was not verified for Fe and Zn in a hydroponic experiment, where lowering Zn and Fe decreased Cd uptake in cacao seedlings (Moore et al., 2024). Field surveys in Latin America showed a near optimal Zn concentration in leaves, suggesting that Zn deficiency is likely not a key factor for Cd uptake (Vanderschueren et al., 2021). In addition, these authors found a strong correlation between leaf and bean Cd, suggesting that leaf analysis can help predict bean Cd. The occurrence of Fe and Mn deficiencies in the field and their possible impact on Cd uptake in cacao is less documented.
Although there is a high variability in the root to shoot transfer of Cd among genotypes, the root to shoot translocation factor ([Cd]shoots/[Cd]roots) is generally > 1 (Galvis et al., 2023; Blommaert et al., 2022, 2025). Branches and stems were identified as major Cd storage compartments, and nibs represented a negligible proportion of total Cd in the tree (Blommaert et al., 2022, 2025).
Concerning the pathways of Cd from roots to nib, a Cd stable isotope tracing experiment suggested that Cd in cacao nibs likely originated from phloem-redistribution from the stem, branches or mature leaves and not from direct root-to-nib transport via the xylem (Vanderschueren et al., 2023). A split-root experiment on cacao seedlings confirmed that Cd was phloem mobile (Blommaert et al., 2024). In mature trees, a progressive retention of Cd compared to Mn and Zn was observed along the transpiration (or xylem) pathway from roots to mature leaves (Blommaert et al., 2025). In particular, Cd was accumulated in phloem rays, phelloderm, and around mucilage cavities in the medulla of branches, after a xylem to phloem transfer along the xylem rays (Blommaert et al., 2022, 2024). The similar Cd isotope signature in branches and nibs suggested that Cd was transferred from branches to nibs instead of being remobilized from mature leaves to nibs (Blommaert et al., 2022, 2025).
5 Cadmium detoxification in the plant
Pot experiments on cacao seedlings grown on Cd-spiked soils at 22.4 to 90 mg Cd kg−1 soil (dos Santos et al., 2020), and 75 to 150 mg Cd kg−1 soil (de Almeida et al., 2022 and 2023) showed strong phytotoxicity symptoms (i.e., Cd damage to membranes and oxidative stress). These above-mentioned studies were conducted in controlled conditions where Cd was added to soils artificially, at unrealistically high Cd concentrations compared to environmental concentrations (Tab. 1). There is no report of visible toxicity symptoms related to Cd in cacao fields, which means that cacao plants cope with Cd toxicity at environmental concentrations. This plant species develops original mechanisms for Cd storage and detoxification. Oxygen ligands are key players in Cd storage in branches and nibs (Blommaert et al., 2022). Calcium oxalate crystals, which are present in all organs (Letort et al, 2025), especially in branches (observed in frozen hydrated state), participate in Cd binding (Blommaert et al., 2024). In the nibs, Cd phytate was suggested as the main Cd species in beans containing 2.5 ± 0.4 mg Cd kg -1. However, a change in Cd speciation to Cd bound to thiol-containing ligands (S-ligands) was observed for Cd-rich beans (9.1 ± 2.6 mg Cd kg -1) (Blommaert et al., 2023), suggesting different Cd detoxification strategies depending on the exposure level. Oxalate is an antinutritional factor, limiting the bioaccessibility of metals. Further studies should be conducted to test the relationship between oxalate concentration and Cd bioaccessibility in chocolate products.
6 Mitigation strategies to reduce cadmium in chocolate
Various mitigation strategies can be applied from harvest to chocolate production to reduce Cd accumulation in cacao (Fig. 3). In this section, each strategy is presented, starting from short to long term ones. We evaluate the results obtained on their efficiency, i.e., whether significant decreases in bean Cd were obtained.
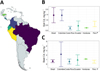 |
Fig. 2 A. Map showing countries with Cd national surveys in Latin America B. Soil Cd for each country (Mean and SD, except for Peru: min and max Cd concentration). C. Bean Cd for each country (Mean and SD, except for Peru: min and max Cd concentration). Data for Brazil do not come from a Nation-scale study, but they were included for comparison. |
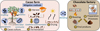 |
Fig. 3 Summary of Cd mitigation methods (in blue) along the chocolate production process, from field to bar. |
6.1 Blending cacao beans from different origins
Because the maximum permitted Cd concentrations defined by the EU and CAC concern chocolate products instead of cacao beans, it is possible for the producers and manufacturers to mix cacao beans from different origins to meet these values in the final products. This practice is forbidden when the MPC is on raw products, as for cereals, vegetables, etc... (EC, 2023/915). It is, of course, a less virtuous strategy than reducing Cd accumulation by the cacao trees. In addition, it involves a loss of traceability and regional identity, and it is not adapted to single origin fine-flavored chocolates, which are mostly produced in Latin America.
6.2 Post-harvest treatments
There is an agreement that testa are richer in Cd than nibs before fermentation (Vanderschueren et al., 2023 and references therein; Bravo et al., 2022). The testa is mostly removed after roasting and before grinding the nibs. The optimization of this removal could help to reduce nib Cd, to a factor 1.16 based on a simple calculation (Vanderschueren et al., 2021). And reduction of Pb is improved by better cleaning of the commercial beans (Ahn et al., 2022).
Several studies hypothesized that fermentation could favour the transfer of Cd from nib to testa. Fermentation is a complex biochemical process leading to bean acidification and chemical transformations. Based on a fermentation experiment in the field, Vanderschueren et al. (2020) suggested that acidification of nib (pH<5) caused migration of Cd by a factor 1.3, from the nibs to the testa. This is valuable to decrease Cd in the final product since most of the testa is removed from the nib after roasting. Vanderschueren et al. (2022) compared lab-scale (5 kg) and field micro-fermentation (>200 kg) of cacao. While artificial acid amendments (lactic and acetic acid) had no effect in lab-scale fermentation, micro-fermentation reduced nib Cd by a factor of 1.25 after four days, suggesting that fermentation enhances Cd mobility. The temperature differences between fermentation volumes would influence acidification-driven Cd mobilization in nibs. Vanderschueren et al. (2023) studied the effects of temperature, acetic acid, and ethanol on cacao in incubation tests mimicking fermentation. While ethanol had no significant impact on nib pH or nib Cd, acetic acid and temperature, combined or not, significantly reduced nib pH and Cd. Under typical fermentation conditions (45°C, 20 g L−1 acetic acid), nib Cd decreased by a factor of 1.3, and under extreme conditions (65°C, 40 g L−1 acetic acid), by a factor of 1.6. By adjusting the fermentation parameters and optimizing the testa removal, there is a possibility to decrease Cd in chocolate. Fermentation is an essential step for developing the aromatic precursors and bioactive compounds characteristic of the aroma in chocolate and cacao products (Koné et al., 2021). Thus, a compromise between low Cd and aroma should be found.
In contrast, Bravo et al. (2022) did not evidence a significant decrease in bean Cd during natural and artificial (incubation with lactic and acetic acid) fermentation.
6.3 Soil amendments
Lime, gypsum, biochar, and Zn supplementation were the first soil amendments investigated for the mitigation of Cd in cacao (Vanderschueren et al., 2021). Thereafter, studies tested the same amendments and others shared new knowledge on compost and Mn supplementation, as described below.
Soil amendment with lime (Arévalo-Hernández et al., 2022) and biochar are known to increase soil pH (Ramtahal et al., 2019). Due to its higher solubility in water compared to lime, gypsum can penetrate deeper soil layers (Liu and Hue, 2001). Argüello et al. (2022) conducted two short-term experiments (pot and incubation) to assess the effect of gypsum on Cd uptake and its influence on Cd phytoavailability. The initial hypothesis was that in Ca-deficient soils, gypsum application would reduce Cd uptake due to competition between Ca2+ and Cd2+. However, gypsum application increased Cd2+ desorption in all soils. Despite this desorption, gypsum did not enhance Cd uptake in cacao seedlings. In two cacao orchards (soil pH 6.6 and 5.1) in Ecuador, Argüello et al. (2023) studied the effects of lime, gypsum, and compost on reducing Cd accumulation over a 30-month period. Lime and compost increased soil pH, while gypsum did not. They significantly reduced leaf Cd but not bean Cd, and were inconsistent over time. A mixture of lime and compost was advocated for future research since compost increases lime penetration into the soil profile, allowing better root growth and stabilisation of the OM. However, liming could cause mineral deficiency. Fertilizers can be used in combination with liming to counter the micronutrients (Zn, Mn, Fe, and Cu) deficiencies caused by liming or overliming (Argüello et al., 2020; Arévalo-Hernández et al., 2022). Villegas et al. (2024) reviewed the use of various biochars (i.e., hardwood, rice husk, corn cob residues, bagasse coffee husk, quinoa straw, and oil palm kernel shell) in different crops (i.e., tomato, cucumber, and cacao), and discussed the advantages and limitations for reducing Cd in cacao in Colombia. Further studies in the field are required to test the efficiency of the various types of biochar.
There is no clear evidence of an amendment or a combination of amendments that effectively reduces Cd accumulation in cacao trees. Because Cd uptake and accumulation in beans can take over 500 days (Vanderschueren et al., 2023), any positive impact of soil amendments may only be observed after a long term, i.e., after 30 months (Argüello et al., 2023). Finally, amendments are often added to mature plantations, which have already bioaccumulated Cd in vegetative parts, especially branches and stems. This Cd stock may contribute to Cd transfer in beans (Blommaert et al., 2022, 2024). This may explain why results in ongoing plantations have not been very effective.
6.4 Bioremediation
Microorganisms could serve as a bioremediation strategy to reduce Cd availability in soil and Cd transfer in cacao. Arce-Inga et al. (2022) selected three Cd-hypertolerant strains of Bacillus (S1C2, R1C2) and of Pseudomonas (V3C3) and reduced the Cd concentration in an agar medium under in vitro conditions. However, in pot experiments, while inoculation with S1C2 and R1C2 showed a reduced Cd concentration in cacao stems, V3C3 did not show any effect on this Cd concentration. Schmidt et al. (2025) identified multiple native strains of Cd-tolerant bacteria and fungi from the cacao rhizosphere by gene sequencing and metabolomics. Among them, the abundance of Paenibacillus sp., and Fusarium sp., known for Cd biosorption, were highly negatively correlated to leaf Cd. Two amplicons from the Ktedonobacterales fungal order were both negatively correlated to leaf and stem Cd (Schmidt et al., 2025). The authors suggest that microbial taxa may be partially responsible for the differentiation in Cd accumulation among cacao genotypes. Further studies are necessary to confirm this hypothesis. Feria-Cáceres et al. (2022) investigated bacterial capacity to immobilize Cd first by laboratory culture in medium, then by inoculation with CCN-51 seeds in greenhouse conditions for bacterial strains isolated from Colombian cacao farm soil. Certain strains such as Exiguobacterium sp., (11-4A), Enterobacter sp., (29-4B) and Klebsiella sp., (18-4B) showed high Cd accumulation and enhanced growth at 3 mg L−1 of Cd. Those strains were selected for inoculation in a pot experiment where seeds were grown with different soil Cd. ANOVA analysis of Cd accumulation found no statistical differences in all treatments.
In conclusion, studies have isolated Cd tolerant microbial strains, which can accumulate Cd in controlled conditions. Some of the reported microorganisms are present in contaminated environments like the Ralstonia sp. At the opposite, some strains were more abundant in the rhizosphere of low Cd accumulating genotypes. These studies open interesting perspectives, but further studies are necessary to assess a significant impact of bacterial inoculation on Cd availability and transfer to cacao beans.
6.5 Cacao genetic diversity and breeding programs
CCN-51 has been widely planted in Latin America because of its high resistance to diseases, its bulk aromatic profile and its adaptability to grow under various environments (full sunlight) (Feria-Cáceres et al., 2022; Jaimez et al., 2022). This clone is a result of several breeding programs aiming to select cultivars based on their yield (Jaimez et al., 2022). In terms of Cd accumulation, this genotype is characterized by a relatively low shoot Cd uptake (see 3.2; Arévalo-Hernández et al., 2021). The variety Nacional is cultivated to produce fine flavour chocolate in Ecuador. Giving Cd accumulation, it was intermediate among the genotypes analyzed by Lewis et al. (2018). Many other cacao cultivars are used, with some regional specificities.
An important variability in bean Cd was found among cultivars (see section 3.2). This variability is an opportunity to select cultivars and/or design new cultivars with low Cd accumulation. To progress in this way, a systematic phenotyping of Cd accumulation in the main genotypes cultivated in Latin America and the Caribbean would be an asset. In addition, knowledge on the ecophysiology of Cd in rice and wheat, and on the transporters and mechanisms involved in Cd uptake and translocation to be transposed in cacao.
Cultivar selection and breeding programs are long term processes. Multiple factors are considered, including yield, resistance to pathogens and stress factors, quality, and Cd accumulation as a new parameter. Cultivar selection can benefit from the large genetic diversity of cacao, which was preserved until now thanks to several cacao genebanks worldwide and to local collections. A challenge of the agrobiodiversity is to preserve it in the future, and to explore the potential of cacao genotypes regarding the multiple factors listed above, using a holistic approach (Lavoie et al., 2023).
6.6 Grafting
Grafting is a common agronomic practice in cacao. In Latin America most cacao trees in orchards are grafted. It can increase productivity (Ocampo-Ariza et al., 2025) and resistance to pathogens (de Almeida et al., 2023). Grafting with a well-chosen combination of rootstock and scion may help for reducing Cd transfer to aboveground parts of cacao trees (Vanderschueren et al., 2021).
Studies from Borjas-Ventura et al. (2022) and Galvis et al. (2023) explored the suitability of genotypes to accumulate Cd in roots and be used as rootstock (see section 3.2). As described in that section, IMC-67 and PA-46 could be used as rootstocks for reducing the translocation of Cd from roots to shoots as they had the lowest Cd accumulation in the plant (Galvis et al., 2023). Fernández-Paz et al. (2021) compared Cd accumulation and toxicity in various rootstocks and grafting combinations, grown on soil with 7.49 mg Cd kg−1, in greenhouse conditions. Significant differences in Cd uptake were found. Difference in Cd tolerance were also suggested. The different PA-121 seedling rootstocks exhibited more toxicity symptoms while grafts with CCN-51 scion showed more stable photosynthetic performance than those with ICS-95. These results must be confirmed with field experiments. de Almeida et al. (2022) tested various grafting combinations using CCN-51, BN-34, PH-16, and PS-13.19. The scion/rootstock combinations CCN-51/BN-34 and CCN-51/PH-16 had a lower Cd translocation to leaves and higher Cd accumulation in roots. Further studies are needed to assess bean Cd across different combinations, in the field.
Low-Cd accumulator genotypes present a promising long-term solution. Further research is needed to understand cacao physiology and accumulation of Cd in grafted plants. In addition, current grafting practices use seeds from open pollination for rootstock propagation, which prevent the conservation of the genotypes. Programs for securing homogeneous genetic material of rootstock propagation should be developed.
6.7 Transfer of knowledge
The knowledge on Cd in cacao described in this review has to be translated into practical guidelines and tools transferred to stakeholders, especially farmers and cacao exporters. Several initiatives have been taken in that direction. A series of 13 guides in Spanish are available online at https://bit.ly/3nreRny. They are accessible via other websites and are easy to find on the web. They propose practical guidelines for soil Cd measurement, risk evaluation and mitigation strategies. In parallel, the Alliance Bioversity-CIAT developed an online tool (https://cacaodiversity.org/) providing information as maps, agroforestry management, and other environmental parameters for cacao plantations at a specific location in Peru. These tools should be maintained and regularly updated with the progress of the knowledge and technologies.
7 Other metallic contaminants in chocolate
Alongside Cd, other worrisome metal(loid)s could occur in cacao beans and therefore merit further discussion. In previous reports, different metals such as nickel (Ni), copper (Cu), Zn, chromium (Cr), Fe, and metalloids, like arsenic (As) were measured in cacao beans and by-products, including chocolate (Ščančar et al., 2013; Romero-Estévez et al., 2019; Barraza et al., 2021; Lewis et al., 2018; Abt and Robin, 2020; Frimpong et al., 2025). The origin of these contaminants is probably multiple. It could (i) be geogenic (ultramafic soils for Ni, for example), (ii) result from local contamination, or (iii) result from post-harvest processing. Recently, the EU adopted MPCs for Ni in chocolate products. They range from 2.5 (milk chocolate with < 30% total dry cocoa solids)) to 7 (milk chocolate with ≥ 30% total dry cocoa solids and chocolate) and 15 (cocoa powder placed on the market) mg Ni kg−1 (EC, 2024/1987). Thus, a chocolate with 30% and 70% of cocoa solids can be made with beans containing up to 8.3 and 10.0 mg Ni kg−1, respectively, supposing the other ingredients contain no Cd. Contrary to Cd, there is no difference in Ni concentration between cacao beans from South America (6.53 ± 2.01 mg Ni kg−1) and from West Africa (average 6.58 ± 1.95 mg Ni kg−1) (Bertoldi et al., 2016). Similarly, Godebo et al. (2024) reported a range of 0.94 - 7.39 and 0.42 - 7.35 mg Ni kg−1 in samples from South America and West Africa, respectively. The values reported in these surveys comply with the new regulation, but more extensive surveys would be necessary since high Ni-containing chocolate products were found in the market a decade ago (Ščančar et al., 2013).
8 Conclusions, perspectives
In recent years, knowledge on Cd transfer in cacao has progressed in many aspects thanks to a large research effort of the community. The number of national surveys of soil and bean Cd has increased, but there are still gaps for a complete view on the contamination pathways. Emerging techniques for Cd analysis in soil and plants such as optimized Monochromatic Energy Dispersive X-ray Fluorescence (MEDXRF) might help to fill the gap and refine the current knowledge. This method is an alternative to Inductively Coupled Plasma - Mass Spectrometry (ICP-MS), with limited sample preparation (grinding and sieving at 500 μm), fast analysis (200s measurement time) and no need of gas and chemicals. It was tested by Dekeyrel et al. (2024) on soil and cacao samples (beans, liquors, and leaves). Strong correlations were found between Cd concentrations measured by MEDXRF and ICP-MS (Cacao bean: R2 = 0.99; Cacao liquor: R2 = 0.98; Cacao leaf: R2 = 0.97; Soil: R2 = 0.99). The average limit of quantification was 0.178 mg Cd kg⁻¹ for plant samples and 0.205 mg Cd kg⁻¹ for soil.
Mitigation strategies are offering perspectives to address the health and socio-economic threat related to Cd in cacao, with short term and long-term options. Post-harvest treatments would be promising. First, processes should target a complete removal of the testa, which is richer in Cd than the nib. Adjusting fermentation parameters should help to decrease nib Cd, although further fine tuning and adaptation to real conditions practical on farms where cacao beans are produced and normally fermented are needed. Concerning soil amendments, evidence of their efficiency in the field is still lacking. Tests could be conducted on young plantations, or with a combination of amendments. Circular economy is a way to explore. For example, Mwafulirwa et al. (2024) found that recycling cacao pod husks (CPH) into biochar, compost, or their mixture can improve nutrient availability in acidic soils by increasing pH. Although this practice may contribute to Cd enrichment in the topsoil like leaf fall (Gramlich et al., 2018), its impact on Cd phytoavailability and potential Cd mitigation could be examined. The benefit of agroforestry for reducing bean Cd is debated, and more studies are necessary to conclude. Agroforestry is beneficial for restoring ecological interactions, decreasing deforestation and increasing the resilience of the ecosystem. However, the decrease of the yield in the short term is an obstacle to its global establishment. Niether et al. (2020) compared 36 studies of both monoculture and agroforestry cacao systems and found an average yield decrease of 25% in agroforestry. However, this decrease may be compensated by a longer lifespan of the orchard. Cultivar selection and grafting are promising due to the large genetic diversity of cacao and variability in Cd accumulation among cultivars. Both rootstock and scion can vary in Cd accumulation and in subsequent transfer to developing seeds. A large screening of cacao genotypes under environmentally realistic conditions is still needed. This is one of the objectives of the international project Climaloca (https://climaloca.org). More generally, it is important to include realistic Cd exposure conditions in experimental designs because results obtained with spiked soils and hydroponics at very high concentrations may not be extrapolated at environmental concentrations. In addition, studies on soil properties (e.g., micronutrients availability in deficit soil) are required to better understand their interaction with Cd and their influence in its accumulation in cacao. Research on Cd mitigation in cacao is young. Although cacao tree differs by many aspects from cereals, this research can be fuelled by the huge knowledge and experience on Cd mitigation in rice and wheat.
In the context of global change and increased anthropogenic pressure, Cd contamination should be considered together with environmental stress parameters impacting cacao plantations. Indeed, plantations are subjected to pests, diseases, infections, and drought, which increase with climate change (Kongor et al., 2024). Globally, a decline in suitable area for cacao cultivation is forecasted in major country producers of West Africa and Latin America (Ceccarelli et al., 2024). Mitigation strategies need to be designed in a holistic approach, in order to make agroecosystems more resilient and sustainable. In order to progress in the design of Cd mitigation strategies, transdisciplinary projects involving researchers and stakeholders are valuable because they engage farmers into field experiments integrating scientific approaches, and include their needs and constraints to the research process.
Cadmium concentrations in mg Cd kg−1 dry weight in cacao farm systems found in national surveys in Latin America.
Acknowledgments
The authors thank both reviewers for their constructive comments and valuable suggestions, which have significantly improved the quality and clarity of this manuscript.
Funding
This work was supported by the French National Research Agency in the framework of the France 2030 investment program ANR-22-EXES-0001 and of the program Labex OSUG ANR-10-LABX56, by the European Synchrotron Radiation Facility (ESRF, Grenoble, France), and by the CNRS/INSU in the framework of the program EC2CO (project CAMINO).
Conflicts of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Data availability statement
The data that support the findings of this article are already openly available.
Author contribution statement
Conceptualization, Formal analysis and Methodology: GS, CC, FL; Investigation and Writing – original draft: GS, CC, EC, FL; Visualization: CC; Validation and Writing – review & editing: All authors. All authors approved the submitted version.
References
- Abt E, Robin LP. Perspective on cadmium and lead in cocoa and chocolate. In: Journal of Agricultural and Food Chemistry. American Chemical Society, 2020, Vol. 68, Issue 46, pp. 13008–13015. https://doi.org/10.1021/acs.jafc.9b08295. [Google Scholar]
- Ahn T, Chaney R, DiBartolomeis M, Ramtahal G. 2022. Expert Investigation Related to Cocoa and Chocolate Products: Final Report. Submitted to: As You Sow and the Settling Defendants. Under order of Superior Court of the State of California, County of San Francisco, Case No. CGC-15-548791. 381 pp. [Google Scholar]
- Araujo Q, Gattward J, Freitas J, Almeida S, Baligar V. 2022. Potentially toxic elements in soils and cacao beans in agroforestry systems of Bahia, Brazil. Agrotrópica (Itabuna) 34(2): 107–120. https://doi.org/10.21757/0103-3816.2022v34n2p107-120. [Google Scholar]
- Arce-Inga M, González-Pérez AR, Hernandez-Diaz E, et al. 2022. Bioremediation potential of native bacillus sp., strains as a sustainable strategy for cadmium accumulation of Theobroma cacao in Amazonas region. Microorganisms 10(11[CE2]). https://doi.org/10.3390/microorganisms10112108. [Google Scholar]
- Arévalo-Gardini E, Arévalo-Hernández CO, Baligar VC, He ZL. 2017. Heavy metal accumulation in leaves and beans of cacao (Theobroma cacao L.) in major cacao growing regions in Peru. Sci Total Environ 605–606: 792–800. https://doi.org/10.1016/j.scitotenv.2017.06.122. [Google Scholar]
- Arévalo-Hernández CO, Arévalo-Gardini E, Barraza F, Farfán A, He Z, Baligar VC. 2021. Growth and nutritional responses of wild and domesticated cacao genotypes to soil Cd stress. Sci Total Environ 763. https://doi.org/10.1016/j.scitotenv.2020.144021. [Google Scholar]
- Arévalo-Hernández CO, Arévalo-Gardini E, Farfan A, et al. 2022. Growth and nutritional responses of juvenile wild and domesticated cacao genotypes to soil acidity. Agronomy 12(12). https://doi.org/10.3390/agronomy12123124. [Google Scholar]
- Argüello D, Chavez E, Gutierrez E, et al. 2023. Soil amendments to reduce cadmium in cacao (Theobroma cacao L.): A comprehensive field study in Ecuador. Chemosphere 324. https://doi.org/10.1016/j.chemosphere.2023.138318. [Google Scholar]
- Argüello D, Chavez E, Lauryssen F, Vanderschueren R, Smolders E, Montalvo D. 2019. Soil properties and agronomic factors affecting cadmium concentrations in cacao beans: A nationwide survey in Ecuador. Sci Total Environ 649: 120–127. https://doi.org/10.1016/j.scitotenv.2018.08.292. [Google Scholar]
- Argüello D, Dekeyrel J, Chavez E, Smolders E. 2022. Gypsum application lowers cadmium uptake in cacao in soils with high cation exchange capacity only: A soil chemical analysis. Eur J Soil Sci 73(2). https://doi.org/10.1111/ejss.13230. [Google Scholar]
- Argüello D, Montalvo D, Blommaert H, Chavez E, Smolders E. 2020. Surface soil liming reduces cadmium uptake in cacao seedlings but subsurface uptake is enhanced. J Environ Qual 49(5): 1359–1369. https://doi.org/10.1002/jeq2.20123. [Google Scholar]
- Barraza F, Schreck E, Lévêque T, et al. 2017. Cadmium bioaccumulation and gastric bioaccessibility in cacao: A field study in areas impacted by oil activities in Ecuador. Environ Pollut 229: 950–963. https://doi.org/10.1016/j.envpol.2017.07.080. [Google Scholar]
- Barraza F, Schreck E, Uzu G, et al. 2021. Beyond cadmium accumulation: Distribution of other trace elements in soils and cacao beans in Ecuador. Environ Res 192. https://doi.org/10.1016/j.envres.2020.110241. [Google Scholar]
- Barroso JP, de Almeida AAF, do Nascimento JL, et al. 2023. The damage caused by Cd toxicity to photosynthesis, cellular ultrastructure, antioxidant metabolism, and gene expression in young cacao plants are mitigated by high Mn doses in soil. Environ Sci Pollut Res Int 30(54): 115646–115665. https://doi.org/10.1007/s11356-023-30561-1. [Google Scholar]
- Bertoldi D, Barbero A, Camin F, Caligiani A, Larcher R. 2016. Multielemental fingerprinting and geographic traceability of Theobroma cacao beans and cocoa products. Food Control 65: 46–53. https://doi.org/10.1016/j.foodcont.2016.01.013. [Google Scholar]
- Blommaert H, Aucour AM, Wiggenhauser M, et al. 2025. Comparison of cadmium pathways in a high Cd accumulating cultivar versus a low Cd accumulating cultivar of Theobroma cacao L. Plant Physiol Biochem 220. https://doi.org/10.1016/j.plaphy.2025.109511. [Google Scholar]
- Blommaert H, Aucour AM, Wiggenhauser M, et al. 2022. From soil to cacao bean: Unravelling the pathways of cadmium translocation in a high Cd accumulating cultivar of Theobroma cacao L. Front Plant Sci 13. https://doi.org/10.3389/fpls.2022.1055912. [Google Scholar]
- Blommaert H, Castillo-Michel H, Veronesi G, et al. 2024. Ca-oxalate crystals are involved in cadmium storage in a high Cd accumulating cultivar of cacao. Environ Exp Bot 221. https://doi.org/10.1016/j.envexpbot.2024.105713. [Google Scholar]
- Blommaert H, De Meese C, Wiggenhauser M, Sarret G, Smolders E. 2024. Evidence of cadmium transport via the phloem in cacao seedlings. Plant Soil. https://doi.org/10.1007/s11104-024-06753-0. [Google Scholar]
- Blommaert H, Sarret G, Chavez E, Smolders E, Vanderschueren R. 2023. Cadmium speciation in cacao beans changes during a fermentation-like incubation. Food Chem 431. https://doi.org/10.1016/j.foodchem.2023.137068. [Google Scholar]
- Borjas-Ventura R, Bello-Medina N, Bello-Amez S, et al. 2022. Differentiated cadmium uptake and its effects on the physiology of six genotypes (Theobroma cacao L.) in San Ramon, Central Peruvian Jungle. Trop Subtrop Agroecosystems 25(3). https://doi.org/10.56369/TSAES.4000. [Google Scholar]
- Bravo D, Araujo-Carrillo G, Carvalho F, et al. 2024. First national mapping of cadmium in cacao beans in Colombia. Sci Total Environ 954. https://doi.org/10.1016/j.scitotenv.2024.176398. [Google Scholar]
- Bravo D, Leon-Moreno C, Martínez CA, et al. 2021. The first national survey of cadmium in cacao farm soil in Colombia. Agronomy, 11(4). https://doi.org/10.3390/agronomy11040761. [Google Scholar]
- Bravo D, Pardo-Díaz S, Benavides-Erazo J, Rengifo-Estrada G, Braissant O, Leon-Moreno C. 2018. Cadmium and cadmium-tolerant soil bacteria in cacao crops from northeastern Colombia. J Appl Microbiol 124(5): 1175–1194. https://doi.org/10.1111/jam.13698. [Google Scholar]
- Bravo D, Santander M, Rodríguez J, Escobar S, Ramtahal G, Atkinson R. 2022. ‘From soil to chocolate bar’: identifying critical steps in the journey of cadmium in a Colombian cacao plantation. Food Addit Contam Part A 39(5): 949–963. https://doi.org/10.1080/19440049.2022.2040747. [Google Scholar]
- Cáceres PFF, Vélez LP, Junca H, Moreno-Herrera CX. 2021. Theobroma cacao L. agricultural soils with natural low and high cadmium (Cd) in Santander (Colombia), contain a persistent shared bacterial composition shaped by multiple soil variables and bacterial isolates highly resistant to Cd concentrations. Curr Res Microbial Sci 2. https://doi.org/10.1016/j.crmicr.2021.100086. [Google Scholar]
- Cadmium dietary exposure in the European population. 2012. In EFSA Journal (Vol. 10, Issue 1). Wiley-Blackwell Publishing Ltd. https://doi.org/10.2903/j.efsa.2012.2551. [Google Scholar]
- Carrillo K, Martínez M, Ramírez L, Argüello D, Chavez E. 2023. Cadmium (Cd) distribution and soil-plant relationship in cacao farms in Costa Rica. Environ Monit Assess 195(10). https://doi.org/10.1007/s10661-023-11817-2. [Google Scholar]
- Ceccarelli V, Fremout T, Chavez E, et al. 2024. Vulnerability to climate change of cultivated and wild cacao in Ecuador. Clim Change 177(7). https://doi.org/10.1007/s10584-024-03756-9. [Google Scholar]
- Clemens S, Ma JF. 2016. Toxic heavy metal and metalloid accumulation in crop plants and foods. In: Annual Review of Plant Biology (Vol. 67, pp. 489–512). Annual Reviews Inc. https://doi.org/10.1146/annurev-arplant-043015-112301. [Google Scholar]
- COMMISSION REGULATION (EU) 2023/915 of 25 April 2023 on maximum levels for certain contaminants in food and repealing Regulation (EC) No 1881/2006. [Google Scholar]
- de Almeida NM, de Almeida AAF, de Almeida Santos N, Mora-Ocampo IY, Pirovani CP. 2023. Leaf proteomic profiles in cacao scion-rootstock combinations tolerant and intolerant to cadmium toxicity. Plant Physiol Biochem 203. https://doi.org/10.1016/j.plaphy.2023.107987. [Google Scholar]
- de Almeida NM, de Almeida AAF, Santos N, et al. 2022. Scion-rootstock interaction and tolerance to cadmium toxicity in juvenile Theobroma cacao plants. Sci Hortic 300. https://doi.org/10.1016/j.scienta.2022.111086. [Google Scholar]
- Dekeyrel J, Atkinson R, Chavez E, et al. 2024. Using optimized monochromatic energy dispersive X-ray fluorescence to determine the cadmium concentration in cacao and soil samples. Heliyon 10(20): e39034. https://doi.org/10.1016/j.heliyon.2024.e39034. [Google Scholar]
- dos Santos MLS, de Almeida AAF, da Silva NM, et al. 2020. Mitigation of cadmium toxicity by zinc in juvenile cacao: Physiological, biochemical, molecular and micromorphological responses. Environ Exp Bot 179. https://doi.org/10.1016/j.envexpbot.2020.104201. [Google Scholar]
- European Commission. 2014. Commission Regulation (EU) No 488/2014 of 12 May 2014 amending Regulation (EC) No 1881/2006 as regards maximum levels of cadmium in foodstuffs. Off J Eur Union 138: 75–79. [Google Scholar]
- Feria-Cáceres PF, Penagos-Velez L, Moreno-Herrera CX. 2022. Tolerance and cadmium (Cd) immobilization by native bacteria isolated in cocoa soils with increased metal content. Microbiol Res 13(3): 556–573. https://doi.org/10.3390/microbiolres13030039. [Google Scholar]
- Fernández-Paz J, Cortés AJ, Hernández-Varela CA, Mejía-de-Tafur MS, Rodriguez-Medina C, Baligar VC. 2021. Rootstock-mediated genetic variance in cadmium uptake by juvenile cacao (Theobroma cacao L.) genotypes, and its effect on growth and physiology. Front Plant Sci 12. https://doi.org/10.3389/fpls.2021.777842 [Google Scholar]
- Frimpong E, Ankapong E, Boakye KO, et al. 2025. Uptake and in-vitro bioaccessibility of toxic metals in cocoa beans: human health risks. Environ Geochem Health 47(2): 33. https://doi.org/10.1007/s10653-024-02345-0. [Google Scholar]
- Galvis DA, Jaimes-Suárez YY, Rojas Molina J, Ruiz R, León-Moreno CE, Carvalho FEL. 2023. Unveiling cacao rootstock-genotypes with potential use in the mitigation of cadmium bioaccumulation. Plants 12(16). https://doi.org/10.3390/plants12162941. [Google Scholar]
- Gil JP, López-Zuleta S, Quiroga-Mateus RY, Benavides-Erazo J, Chaali N, Bravo D. 2022. Cadmium distribution in soils, soil litter and cacao beans: a case study from Colombia. Int J Environ Sci Technol 19(4): 2455–2476. https://doi.org/10.1007/s13762-021-03299-x. [Google Scholar]
- Godebo TR, Stoner H, Kodsup P, et al. 2024. Occurrence of heavy metals coupled with elevated levels of essential elements in chocolates: Health risk assessment. Food Res Int 187. https://doi.org/10.1016/j.foodres.2024.114360. [Google Scholar]
- Gramlich A, Tandy S, Andres C, et al. 2017. Cadmium uptake by cocoa trees in agroforestry and monoculture systems under conventional and organic management. Sci Total Environ 580: 677–686. https://doi.org/10.1016/j.scitotenv.2016.12.014 [Google Scholar]
- Gramlich A, Tandy S, Gauggel C, et al. 2018. Soil cadmium uptake by cocoa in Honduras. Sci Total Environ 612: 370–378. https://doi.org/10.1016/j.scitotenv.2017.08.145. [Google Scholar]
- Guarín D, Hamamura NR, Cortez JB, et al. 2023. Cadmium contamination in cacao farms of Piura, North Peru: A comprehensive assessment of geogenic and anthropogenic sources and implications for future production. Environ Chall 13. https://doi.org/10.1016/j.envc.2023.100765. [Google Scholar]
- Guarín D, Martín-López JM, Libohova Z, et al. 2024. Accumulation of cadmium in soils, litter and leaves in cacao farms in the North Sierra Nevada de Santa Marta, Colombia. Geoderma Reg 36. https://doi.org/10.1016/j.geodrs.2024.e00762. [Google Scholar]
- Gutiérrez E, Chávez E, Gamage KHH, Argüello D, Galkaduwa MB, Hettiarachchi GM. 2022. Cadmium fractionation in soils affected by organic matter application: Transfer of cadmium to cacao (Theobroma cacao L.) tissues. Front Environ Sci 10. https://doi.org/10.3389/fenvs.2022.954521. [Google Scholar]
- Jaimez RE, Barragan L, Fernández-Niño M, et al. 2022. Theobroma cacao L. cultivar CCN 51: a comprehensive review on origin, genetics, sensory properties, production dynamics, and physiological aspects. PeerJ 9. PeerJ Inc. https://doi.org/10.7717/peerj.12676. [Google Scholar]
- Jaramillo-Mazo C, Bravo D, Guerra Sierra BE, Alvarez JC. 2024. Association between bacterial community and cadmium distribution across Colombian cacao crops. Microbiol Spectr 12(7). https://doi.org/10.1128/spectrum.03363-23. [Google Scholar]
- Joya-Barrero V, Huguet C, Pearse J. 2023. Natural and anthropogenic sources of cadmium in cacao crop soils of Santander, Colombia. Soil Syst 7(1). https://doi.org/10.3390/soilsystems7010012. [Google Scholar]
- Koné KM, Assi-Clair BJ. Kouassi ADD, et al. 2021. Pod storage time and spontaneous fermentation treatments and their impact on the generation of cocoa flavour precursor compounds. Int J Food Sci Technol 56(5): 2516–2529. https://doi.org/10.1111/ijfs.14890. [Google Scholar]
- Kongor JE, Owusu M, Oduro-Yeboah C. 2024. Cocoa production in the 2020s: challenges and solutions. In: CABI Agriculture and Bioscience (Vol. 5, Issue 1). BioMed Central Ltd. https://doi.org/10.1186/s43170-024-00310-6. [Google Scholar]
- Lavoie A, Thomas E, Olivier A. 2023. Local working collections as the foundation for an integrated conservation of Theobroma cacao L. in Latin America. Front Ecol Evol 10. https://doi.org/10.3389/fevo.2022.1063266. [Google Scholar]
- Letort F, Chavez E, Blommaert H, et al. 2025. Calcium oxalate crystals in cacao trees and their interactions with cadmium. Plant Physiol Biochem 220. https://doi.org/10.1016/j.plaphy.2025.109499. [Google Scholar]
- Lewis C, Lennon AM, Eudoxie G, Umaharan P. 2018. Genetic variation in bioaccumulation and partitioning of cadmium in Theobroma cacao L. Sci Total Environ 640–641: 696–703. https://doi.org/10.1016/j.scitotenv.2018.05.365. [Google Scholar]
- Liu J, Hue NV. 2001. Amending subsoil acidity by surface applications of gypsum, lime, and composts. Commun Soil Sci Plant Anal 32(13–14): 2117–2132. https://doi.org/10.1081/CSS-120000273. [Google Scholar]
- Maddela NR, Kakarla D, García LC, Chakraborty S, Venkateswarlu K, Megharaj M. 2020. Cocoa-laden cadmium threatens human health and cacao economy: A critical view. In: Science of the Total Environment (Vol. 720). Elsevier B.V. https://doi.org/10.1016/j.scitotenv.2020.137645. [Google Scholar]
- McLaughlin MJ, Smolders E, Zhao FJ, Grant C, Montalvo D. 2021. Managing cadmium in agricultural systems. In: Advances in Agronomy (Vol. 166, pp. 1–129). Academic Press Inc. https://doi.org/10.1016/bs.agron.2020.10.004. [Google Scholar]
- Meter A, Atkinson RJ, Laliberte B. 2019. Cadmium in cacao from Latin America and the Caribbean – A review of research and potential mitigation solutions. Bioversity International, Rome, October 2019. Retrieved from https://scioteca.caf.com/handle/123456789/1506. [Google Scholar]
- Moore RET, Ullah I, de Oliveira VH, et al. 2020. Cadmium isotope fractionation reveals genetic variation in Cd uptake and translocation by Theobroma cacao and role of natural resistance-associated macrophage protein 5 and heavy metal ATPase-family transporters. Hortic Res 7(1). https://doi.org/10.1038/s41438-020-0292-6. [Google Scholar]
- Moore RET, Ullah I, Dunwell JM, Rehkämper M. 2024. Stable isotope analyses reveal impact of Fe and Zn on Cd uptake and translocation by Theobroma cacao. Plants 13(4). https://doi.org/10.3390/plants13040551. [Google Scholar]
- Mwafulirwa L, Sizmur T, Daymond A, et al. 2024. Cocoa pod husk-derived organic soil amendments differentially affect soil fertility, nutrient leaching, and greenhouse gas emissions in cocoa soils. J Clean Prod 479. https://doi.org/10.1016/j.jclepro.2024.144065. [Google Scholar]
- Niether W, Jacobi J, Blaser WJ, Andres C, Armengot L. 2020. Cocoa agroforestry systems versus monocultures: A multi-dimensional meta-analysis. Environ Res Lett 1 5(10). https://doi.org/10.1088/1748-9326/abb053. [Google Scholar]
- Ocampo-Ariza C, Müller S, Yovera F, et al. 2025. Cacao grafting increases crop yield without compromising biodiversity. J Appl Ecol. https://doi.org/10.1111/1365-2664.14851. [Google Scholar]
- Olego MÁ, Quiroga MJ, Sánchez-García M, Cuesta M, Cara-Jiménez J, Garzón-Jimeno JE. 2021. Effects of overliming on the nutritional status of grapevines with special reference to micronutrient content. Int Vitic Enol Soc. https://doi.org/10.20870/oeno-one.2021.55.2.4081. [Google Scholar]
- Ortiz-Álvarez A, Magnitskiy S, Silva-Arero EA, Rodríguez-Medina C, Argout X, Castaño-Marín ÁM. 2023. Cadmium accumulation in cacao plants (Theobroma cacao L.) under drought stress. Agronomy 13(10). https://doi.org/10.3390/agronomy13102490. [Google Scholar]
- Ramtahal G, Umaharan P, Hanuman A, Davis C, Ali L. 2019. The effectiveness of soil amendments, biochar and lime, in mitigating cadmium bioaccumulation in Theobroma cacao L. Sci Total Environ 693. https://doi.org/10.1016/j.scitotenv.2019.07.369. [Google Scholar]
- Romero-Estévez D, Yánez-Jácome GS, Simbaña-Farinango K, Navarrete H. 2019. Content and the relationship between cadmium, nickel, and lead concentrations in Ecuadorian cocoa beans from nine provinces. Food Control 106. https://doi.org/10.1016/j.foodcont.2019.106750. [Google Scholar]
- Ščančar J, Zuliani T, Milačič R. 2013. Study of nickel content in Ni-rich food products in Slovenia. J Food Compos Anal 32(1): 83–89. https://doi.org/10.1016/j.jfca.2013.06.011. [Google Scholar]
- Schmidt JE, Lewis CA, Firl AJ, Umaharan P. 2025. Microbial bioindicators associated with cadmium uptake in sixteen genotypes of Theobroma cacao. Heliyon 11(2). https://doi.org/10.1016/j.heliyon.2025.e41890. [Google Scholar]
- Smolders E, Mertens J. 2013. ‘Cadmium’, in heavy metals in soils: trace metals and metalloids in soils and their bioavailability, 3rd edn. Dordrecht: Springer, pp. 283–308. Available at: https://doi.org/10.1007/978-94-007-4470-7. [Google Scholar]
- Sterckeman T, Thomine S. 2020. Mechanisms of cadmium accumulation in plants. Crit Rev Plant Sci 39(4): 322–359. https://doi.org/10.1080/07352689.2020.1792179. [Google Scholar]
- Thomas E, Atkinson R, Zavaleta D, Rodriguez C, Lastra S, Yovera F, Arango K, Pezo A, Aguilar J, Tames M, Ramos A, Cruz W, Cosme R, Espinoza E, Chavez CR, Ladd B. 2023. The distribution of cadmium in soil and cacao beans in Peru. Sci Total Environ 881. https://doi.org/10.1016/j.scitotenv.2023.163372. [Google Scholar]
- Ullah I, Wang Y, Eide DJ, Dunwell JM. 2018. Evolution, and functional analysis of Natural Resistance-Associated Macrophage Proteins (NRAMPs) from Theobroma cacao and their role in cadmium accumulation. Sci Rep 8(1). https://doi.org/10.1038/s41598-018-32819-y. [Google Scholar]
- Vallejos-Torres G, Gaona-Jimenez N, Arevalo AA, et al. 2023. Cadmium uptake and mycorrhization by cacao clones in agroforestry and monoculture systems of Peruvian amazon. Bioagro 35(3): 237–246. https://doi.org/10.51372/bioagro353.7. [Google Scholar]
- Vanderschueren R, Argüello D, Blommaert H, et al. 2021. Mitigating the level of cadmium in cacao products: Reviewing the transfer of cadmium from soil to chocolate bar. In: Science of the Total Environment (Vol. 781). Elsevier B.V. https://doi.org/10.1016/j.scitotenv.2021.146779. [Google Scholar]
- Vanderschueren R, De Mesmaeker V, Mounicou S, et al. 2020. The impact of fermentation on the distribution of cadmium in cacao beans. Food Res Int 127. https://doi.org/10.1016/j.foodres.2019.108743. [Google Scholar]
- Vanderschueren R, Doevenspeck J, Goethals L, Andjelkovic M, Waegeneers N, Smolders E. 2023. The contribution of cacao consumption to the bioaccessible dietary cadmium exposure in the Belgian population. Food Chem Toxicol 172. https://doi.org/10.1016/j.fct.2023.113599. [Google Scholar]
- Vanderschueren R, Doevenspeck J, Helsen F, et al. 2022. Cadmium migration from nib to testa during cacao fermentation is driven by nib acidification. LWT 157. https://doi.org/10.1016/j.lwt.2022.113077. [Google Scholar]
- Vanderschueren R, Helsen F, Doevenspeck J, Delcour JA, Smolders E. 2023. Incubation tests mimicking fermentation reveal that phytate breakdown is key to lower the cadmium concentrations in cacao nibs. Food Chem 398. https://doi.org/10.1016/j.foodchem.2022.133899. [Google Scholar]
- Vanderschueren R, Wantiez L, Blommaert H, Flores J, Chavez E, Smolders E. 2023. Revealing the pathways of cadmium uptake and translocation in cacao trees (Theobroma cacao L.): A 108Cd pulse-chase experiment. Sci Total Environ 869. https://doi.org/10.1016/j.scitotenv.2023.161816. [Google Scholar]
- Vázquez-deCastro JL, Chávez E, Espinel R, Hendrix S, Smolders E, Maertens M. 2024. The economic costs of cadmium non-tariff measures for smallholder cocoa farmers in Ecuador. World Econ 47(3): 1147–1166. https://doi.org/10.1111/twec.13488. [Google Scholar]
- Villegas CL, Quiroga NJ, Dávalos A, Pizano C. 2024. Reducing cadmium in cocoa (Theobroma cacao) cultivars: analysis of advantages and limitations of different strategies and their potential application in Colombia. Int J Agron 2024(1). https://doi.org/10.1155/ioa/5342167. [Google Scholar]
- Wade J, Ac-Pangan M, Favoretto VR, Taylor AJ, Engeseth N, Margenot AJ. 2022. Drivers of cadmium accumulation in Theobroma cacao L. beans: A quantitative synthesis of soil-plant relationships across the Cacao Belt. PLoS ONE 17(2 February). https://doi.org/10.1371/journal.pone.0261989. [Google Scholar]
- WHO/IARC. 2025. International agency for research on cancer IARC. Monographs on the evaluation of carcinogenic risks for human. Available online at: https://monographs.iarc.fr/agents-classified-by-the-iarc/ (accessed May, 26, 2025). [Google Scholar]
Cite this article as: Letort F, Chavez E, Cesaroni C, Castillo-Michel H, Sarret G. 2025. Cadmium and other metallic contaminants in cacao: Update on current knowledge and mitigation strategies. 32: 22. https://doi.org/10.1051/ocl/2025019
All Tables
Cadmium concentrations in mg Cd kg−1 dry weight in cacao farm systems found in national surveys in Latin America.
All Figures
|
Fig. 1 Production of cacao beans per country in tons in 2022. Only countries ≥ 800 tons were taken into consideration (FAOSTAT, 2022). |
| In the text | |
|
Fig. 2 A. Map showing countries with Cd national surveys in Latin America B. Soil Cd for each country (Mean and SD, except for Peru: min and max Cd concentration). C. Bean Cd for each country (Mean and SD, except for Peru: min and max Cd concentration). Data for Brazil do not come from a Nation-scale study, but they were included for comparison. |
| In the text | |
|
Fig. 3 Summary of Cd mitigation methods (in blue) along the chocolate production process, from field to bar. |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.