")
")
| Issue |
OCL
Volume 31, 2024
Rapeseed / Colza
|
|
|---|---|---|
| Article Number | 5 | |
| Number of page(s) | 12 | |
| Section | Agronomy | |
| DOI | https://doi.org/10.1051/ocl/2024004 | |
| Published online | 09 April 2024 | |
Research article
Chemical composition and quality of rapeseed meal as affected by genotype and nitrogen fertilization☆
Effet de la fertilisation azotée et du génotype sur la composition chimique et la qualité des tourteaux chez le colza
1
Research Unit of Agronomy and Plant Physiology, Regional Agricultural Research Center of Meknes, National Institute of Agricultural Research, PO Box 415, Rabat 10090, Morocco
2
Laboratory of Biotechnologies and Valorization of Bio-Resources (BioVaR), Department of Biology, Faculty of Science, Moulay Ismail University of Meknes, PO Box 11201 Zitoune Meknes 50100, Morocco
3
Department of Animal Production and Pastoralism, National School of Agriculture, PO Box S/40, Meknes 50000, Morocco
4
Research Unit of Plant Breeding and Plant Genetic Resources Conservation, Regional Agricultural Research Center of Meknes, National Institute of Agricultural Research, PO Box 415, Rabat 10090, Morocco
5
Department of Agronomy and Plant Breeding, National School of Agriculture, PO. Box S/40, Meknes 50000, Morocco
* Correspondence: This email address is being protected from spambots. You need JavaScript enabled to view it.
; This email address is being protected from spambots. You need JavaScript enabled to view it.
Received:
10
July
2023
Accepted:
6
February
2024
Abstract
Rapeseed meal (RSM) is known for its high nutritional quality as animal feed. However, there has been little studies on the effect of nitrogen fertilization on RSM chemical composition, mainly neutral detergent fiber (NDF), acid detergent fiber (ADF), acid detergent lignin (ADL), and ash content. Therefore, this study was carried out to evaluate the effect of nitrogen application rate on chemical composition of RSM in interaction with different spring rapeseed varieties (Brassica napus L). A field experiment was conducted during 2018/19 cropping season at the experimental station of the Ecole Nationale d’Agriculture de Meknès” according to a split-plot design with three replications, using six nitrogen application rates, as main plot, and six rapeseed varieties, as subplot. After seeds harvest and oil extraction, meals derived from the different treatments were used in this study. Results showed that increasing nitrogen rate from 0 to 120 kg N ha−1 led to a significant rise in meal yield up to 74.58%. A positive effect of nitrogen fertilization was observed on dry matter, protein content and ash content, recording the highest values at 120 kg N ha−1 treatment. However, cellulose and lignin content were affected negatively by nitrogen fertilization. The nitrogen supply of 150 kg N ha−1 resulted in a reduction of ADF and ADL contents by 23% and 28%, respectively, compared to the unfertilized control (N0). Variability within rapeseed varieties for all parameters except dry matter and ADL content was highlighted.
Résumé
Le tourteau du colza (RSM) est reconnu pour sa meilleure qualité nutritionnelle en tant qu’aliment de bétail. Cependant, les études de l’effet de la fertilisation azotée sur sa composition chimique, notamment les fibres détergentes neutres (NDF), les fibres détergentes acides (ADF), la lignine détergente acide (ADL) et la teneur en cendres, sont rares. Ainsi, cette étude a été menée pour évaluer l’effet de la dose d’azote appliquée sur la composition chimique du RSM en interaction avec différentes variétés de colza de printemps (Brassica napus L). Un essai au champ a été alors conduit durant la saison agricole 2018/2019 à la station expérimentale de l’École Nationale d’Agriculture de Meknès et a concerné six variétés de colza, à savoir Narjisse, Moufida, Alia, Adila, Lila et Trapper, en interaction avec six apports d’azote, soient 0, 30, 60, 90, 120 et 150 kg N ha−1, désignant les traitements respectifs de N0, N30, N60, N90, N120 et N150. Le dispositif expérimental adopté est un split-plot avec trois répétitions, où le facteur azote est attribué à la grande parcelle, alors que la variété est affectée à la petite parcelle. Après la récolte et l’extraction de l’huile, les tourteaux issus des différents traitements ont été utilisés dans cette étude. Les résultats ont montré que l’augmentation de l’apport d’azote de 0 à 120 kg N ha−1 a permis une amélioration significative du rendement en tourteau allant jusqu’à 74,58%. Un effet positif de la fertilisation azotée a été également observé sur la teneur en matière sèche, la teneur en protéines et la teneur en cendres, dont les valeurs les plus élevées ont été enregistrées pour le traitement N120. Cependant, la teneur en cellulose et la teneur en lignine ont été affectées négativement par cette fertilisation. Le traitement N150 a entraîné une réduction des teneurs en ADF et en ADL de 23 et 28%, respectivement, par rapport au témoin non fertilisé (N0). Une variabilité génotypique a été mise en évidence pour tous les paramètres, à l’exception de la matière sèche et de la teneur en ADL.
Key words: rapeseed meal / fertilization / nitrogen / variety / chemical composition
Mots clés : Tourteau de colza / Fertilisation / Azote / Variété / Composition chimique
Contribution to the Topical Issue: “Rapeseed / Colza”.
© M. Yahbi et al., Published by EDP Sciences, 2024
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Highlight
Nitrogen fertilization (NF) significantly affect the rapeseed meal components and quality. It seems that NF improves the meal quality of rapeseed by improving meal yield, protein content, dry matter, and ash content, while reducing fiber content (ADF and ADL).
Nitrogen rate of 120 kg N ha-1 could be the optimum nitrogen rate to produce high meal quality in Moroccan conditions.
For all N rates combined, genotypic variations were found for all traits investigated. Moreover, Trapper the hybrid variety has recorded the highest meal and protein yields, which could be recommended under a nitrogen rate of 120 kg N ha-1.
Introduction
Rapeseed (Brassica napus L.) is one of the most important oilseed crops in the world. It holds third place after palm oil and soybean as a source of vegetable oil. Rapeseed also provides a source of material rich in vegetable protein (meal), which can be included in animal feed, mainly for ruminants but also for pigs, fish, and to a lesser extent for poultry (Peyronnet et al., 2010; Hannoufa et al., 2014; Sanjayan et al., 2014). Indeed, studies on the chemical composition of RSM highlight the relatively high protein content compared to the meals of other major oilseed crops, except soybean (Enami, 2011). Oil-free rapeseed meal contains 35–40% crude protein (Nesi et al., 2008; Sanjayan et al., 2014; Cheng et al., 2022). Although it is comparable in protein with soybean meal, but due to the presence of antinutritional factors including fiber content it has a generally lower digestibility (Khajali and Slominski, 2012).
However, the large-scale incorporation of Rapeseed protein into livestock feed is hampered by the presence of high levels of antinutrients, such as glucosinolates, sinapine, phytates, and fiber (Bellostas et al., 2007; Hannoufa et al., 2014).
The dietary fiber content of rapeseed meal is high compared to soybeans, the cotyledon fiber has a higher negative effect on digestibility of proteins than fibre isolated from seed coat (Bjergegaard et al., 1991). Fiber is among the most important traits that determines the quality and palatability of rapeseed meal. A high fiber content can have detrimental effects on the digestibility and feed efficiency of livestock (Bellostas et al., 2007). Dietary fiber accounts for almost a third of the remaining RSM after oil extraction. It was reported that rapeseed types differed in fiber content. Ochodzki et al. (1995) have reported levels of 27.5–33.0% insoluble dietary fiber and 3.1–5.2% soluble dietary fiber for spring and winter type rapeseed. Moreover, Slominski et al. (2012) concluded that the new yellow-seeded B. napus canola appears to have superior quality characteristics compared with that of its black-seeded counterpart as manifested by its higher protein and sucrose (energy) contents and lower dietary fiber content. Canola-quality Brassica juncea canola has reduced concentrations of protein, sucrose, and dietary fiber and a potentially lower nutritive value compared with that of yellow-seeded B. napus.
Fibers consist of neutral detergent fiber (NDF) comprises hemicellulose, cellulose, lignin and a part of the pectins, acid detergent fiber (ADF) contains cellulose and lignin, and acid detergent lignin (ADL) represents the non-digestible lignin fraction (Van Soest et al., 1991). At our knowledge a few studies have investigated to study the effect of nitrogen level on fiber content in rapeseed (Ribaillieret et al., 1995; Costa et al. 2013). Costa et al. (2013) reported that the nitrogen fertilization affects negatively the ADF and ADL content, but not the NDF content. All these fibers are mainly located in the seed coat (Matthäus, 2002; Wittkop et al., 2009). Indigestible lignin constitute a considerable part of the fibers (Bell, 1995). Lignin is one of most important compositional factors affecting feed utilisation by ruminant. It has a strong effect on the rumen degradability of cellulose and its presence limits the energy value of the meal. Compared to other crops, B. napus contains large amounts of lignin (8% of meal component), whereas in soybean meal it constitutes only 1%.
Since the 80s of the last century, much efforts have been made in research, breeding, and genetic engineering to improve the composition of RSM by reducing levels of anti-nutritional factors and enhancing levels of favourable metabolites (Badani et al., 2005; Hüsken et al., 2005; Weier et al., 2007; Hasan et al., 2008; Nesi et al., 2008; Slominski et al., 2012; Ton et al., 2020). At the beginning researchers were focused mainly on the reduction of glucosinolate levels until the end of the 1980s. Since then, selection has been more limited and has mainly focused on reducing fibre content in favour of protein content, with little effect on the composition of industrial meal. Thus, the main components of rapeseed meal have been reviewed with emphasis on the methods of improving the nutritional value (Cheng et al., 2022). However, the chemical composition of RSM can be changed between rapeseed and canola type (Tab. 1).
Many factors can influence the protein content of rapeseed meal, mainly, the variety, the culture medium, the fiber content, the climatic conditions, and the level of available nitrogen (Hossain et al., 2019; Watts et al., 2021; Cheng et al., 2022). Agronomic practices can so be exploited to improve the protein content and quality of seeds (Kenney et al., 2021).
Researchers have reported variability within rapeseed varieties both for the seed protein content (Olukosi et al., 2017; Watts et al., 2021) and composition (Cruciferin/napin ratio) (Raab et al., 1992; Malabat et al., 2003). Cruciferin (12S globulin) and napin (2S albumin) are storage protein (Schatzki et al., 2014). Both represent up to 70% of seed protein (Nesi et al., 2008). Kasperzak et al. (2016) reported that the content of amino acid and crud protein in RSM and rapeseed cake were significantly affected by the rapeseed variety.
It was reported that climatic conditions such as water and temperature stresses impact B. 100 Napus protein (Hammac et al., 2017). Atmospheric temperature stress has a positive effect on rapeseed protein. Increasing temperature stress increase significantly crude protein content of oilseed rape (Aksouh-Harradj et al., 2006; Hammac et al., 2017). Whereas there is an optimum level of water stress for a maximum crude protein content (Hammac et al., 2017).
Many authors have mentioned a positive effect of nitrogen fertilization on the protein content in rapeseed meal, often associated with a decrease in seed oil content (Brennan and Bolland, 2007; Ahmad et al., 2009; Aminpanah, 2013; Zuo et al., 2016; Hossain et al., 2019). The increase in protein content with nitrogen application can exceed 20% (Zuo et al., 2016) and gradually improves with increasing nitrogen levels (Ahmad et al., 2009). The high protein content at high N level may be due to the negative correlation between oil content and protein content (Hao et al., 2004; Zhao et al., 2006), as oil content generally decreases with increasing N level (Ahmad et al., 2009). Indeed, nitrogen intensifies the synthesis of proteins to the detriment of the of fatty acids synthesis, leading to a reduction in the oil content of the seed (Rathke et al., 2005). The effects of nitrogen on protein content are not always consistent as recorded by Hammac et al. (2017). In their research, the effect of N varied by site and year. However, in all cases where there was a significant effect, the effect was positively related to N dose.
Likewise, little research has been carried out to investigate the genotype for Fiber content and composition (NDF, ADF, ADL and ash) (Kasperzak et al., 2016; Watts et al., 2021). Kasperzak et al. (2016) showed that varietal selection of rapeseed associated with an appropriate extraction method could lead to an improvement in the digestibility of amino acids and crude protein in the RSM.
In Morocco, no study was caried on Moroccan rapeseed varieties concerning the effect of nitrogen fertilization or genotypic variation on meal production and chemical composition. So, the objective of this study was to assess the effect of nitrogen application level and genotype on the chemical composition of RSM from six Moroccan varieties grown under Moroccan rainfed conditions.
Chemical composition of rapeseed and canola meal.
2 Materials and methods
2.1 Plant materiel and field experiment
Six rapeseed varieties, including five Moroccan inbred lines, namely “Narjisse”, “Moufida”, “Alia”, “Adila” and “Lila”, (These are the first Moroccan rapeseed varieties to be registered in the country’s official catalogue and adapted to the local conditions) and a German hybrid, “Trapper” are used in this study. All these cultivars were grown under six nitrogen levels: 0, 30, 60, 90, 120, and 150 kg N ha−1 corresponding, respectively, to N0 (without nitrogen application), N30 (30 kg N ha−1applied at sowing), N60 (split as 30 kg N ha−1 at sowing and 30 kg N ha−1 at rosette stage), N90 (split as 30 kg N ha−1 at sowing, 30 kg N ha−1 at rosette stage and 30 kg N ha−1 at initial flowering), N120 (split as 30 kg N ha−1 at sowing, 60 kg N ha−1 at rosette stage and 30 kg N ha−1 at initial flowering) and N150 (split as 30 kg N ha−1 at sowing, 60 kg N ha−1 at rosette stage and 60 kg N ha−1 at initial flowering). Nitrogen applications were applied manually by the same person using ammonium nitrate (33.5%) fertilizer. The experiment was conducted at the pedagogic farm of the Ecole Nationale d’Agriculture de Meknes (ENAM), Morocco (33°84’ N, 5°47’ W), during 2018/19 cropping season, following a split-plot design with three replications, where the nitrogen treatment was affected to the main plot and the variety to the sub-plot. Each variety was planted in three 5 m long rows spaced 60 cm apart. At harvest stage, one meter from the central raw of each variety from each nitrogen treatment was harvested and threshed. Seed samples from the three replicates were taken and analyzed separately. The soil was a clay loam of the calcimagnesic soil class. At each cropping season, and before sowing, soil samples were taken from the research plot at a depth of 30 cm for organic and chemical analysis (Tab. 2). Planting was done by handplanted on 02 December 2018, the biggening of flowering was at 02 April 2019 and the samples were harvested at 23 May 2019. During the production cycle there was no pressure of pathogens except weed, and the weed management was carried out by hand weeding when it’s necessary. After harvest the seed samples were dried at the ambient temperature for few days and stored at the storage cold room with low moisture.
Rapeseed oil for each seed sample was extracted using the Soxhlet apparatus, model SER 148/6 (VELP SCIENTIFICA, Usmate Velate, Italy). Twenty (20) g of ground seeds (crushed before by a grinder KM-100-CUP) was mixed with 150 mL of hexane for 3 h at 130 °C. The remaining product (rapeseed meal), obtained after oil extraction, was used for chemical analysis. The meal yield was calculated by multiplying by-product content by seed yield (MY, kg ha−1).
Soil characteristics before sowing for year 1 and year 2 cropping seasons.
2.2 Analytical procedures
All analytical procedures were performed at the laboratory of the Department of Animal Production and Pastoralism of Ecole Nationale d’Agriculture de Meknes, Morocco.
Dry matter content in rapeseed meal samples was determined by oven drying at 103 °C for 24 h, whereas ash content was obtained by incineration at 650 °C for 8 h. Total nitrogen (N) was determined following the Kjeldahl method using selenium as catalyst and a K-350 distillation unit (Buchi). Nitrogen content was converted into protein by multiplying by the coefficient 6.25. Neutral detergent fiber (NDF), acid detergent fiber (ADF) and acid detergent lignin (ADL) were measured using an ANKOM 220 Fiber Analyser (Ankom Technology), as described by Van Soest et al. (1991).
In short NDF, ADF and ADL contents were determined in 500 mg meal samples. An Ankom system detergent fiber analysis system with filter bag technology using (Ankom 200, fiber analyser; ANKOM Technology) and heat-stable α-amylase without sodium sulfate. And the NDF, ADF and ADL content were calculated following the equations:
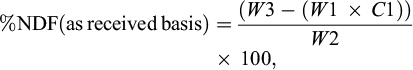 (1)
(1)
where: W1 is the bag tare weight, W2 is the sample weight, W3 is the dried weight of bag with fiber after extraction process, C1 is the blank bag correction (running average of final oven-dried weight divided by original blank bag weight).
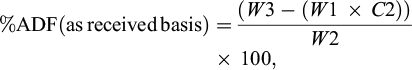 (2)
(2)
where: W1 is the bag tare weight, W2 is the sample weight, W3 is the dried weight of bag with fiber after extraction process, C1 is the blank bag correction (running average of final oven-dried weight divided by original blank bag weight).
 (3)
(3)
where: W1 is the bag tare weight, W2 is the sample weight, W3 is the dried weight of bag with fiber after extraction process, C1 is the blank bag correction (running average of final oven-dried weight divided by original blank bag weight).
2.3 Statistical analysis
Data gathered were subjected to analysis of variance and analysis of correlation using the general linear model (GLM) and correlation procedures, respectively, of the software SPSS for Windows (Version 26). In this study, all factors investigated were considered as fixed. In case of significant differences among varieties or nitrogen treatments, Duncan’s new multiple range test was applied to compare their respective means at 5% probability level.
3 Results
3.1 Weather conditions
Monthly rainfall and air temperature of the cropping season are shown in Figure 1. Recorded rainfall was 224 mm. There was a water deficit due to low precipitation in December and to overcome water deficit, a supplementary irrigation of 50 mm has been provided. This was done at the seedling stage.
 |
Fig. 1 Monthly precipitations (bars) and air temperatures (lines) recorded in the experimental station during the cropping season. |
3.2 Meal yield
The effect of nitrogen application rate and variety was significant on meal yield; however, that of their interaction was not (Tab. 3). For all varieties together, the meal yield rose up significantly with approximately an average of 74.6% (from 1023 to 1786 kg ha−1) by increasing N rate from N0 to N120. Meal yield (MY) variation in the different rapeseed varieties according to the nitrogen application rate in Figure 2.
All the varieties investigated reached their maximum at N120 treatment under which the highest yield was 2019 kg ha−1, recorded in Trapper, followed by 1973 kg ha−1, observed in Alia, while the lowest values were 1467 and 1605 kg ha−1, displayed in Lila and Adila, respectively. In absence of any nitrogen fertilization (N0), once again, Trapper registered the highest MY (1344 kg ha−1) whereas Lila recorded the lowest one (740 kg ha−1).
Main effect of nitrogen rate and variety and their interaction on meal yield and its chemical proprieties of RSM as fed (%). (Data under the nitrogen fertilizer treatment were averaged across varieties, and data under the variety treatment are averaged across fertilizer levels.)
 |
Fig. 2 Meal yield in six rapeseed varieties as affected by nitrogen rate. |
3.3 Protein content
The effect of nitrogen application level and variety was highly significant on protein content (PC). On the other hand, no significant effect on this trait was found for their interaction (Tab. 3). As shown in Figure 3, elevating nitrogen application leads to the increase of PC in all the varieties investigated to reach their maximum at N120, except for Adila which reached its maximum value at N150 treatment.
For all the six varieties together, PC rose up from 36.2% for control (N0) to 44.9%, obtained for N120 treatment, which was not statistically different from 44.5%, observed for N150 treatment. Regarding the varietal variation, and for all the nitrogen rates combined, the highest PC was recorded in Adila, with a mean value of 44.1%, which is statistically comparable to Trapper, with an average of 42.9%. Whereas the lowest PC was 39.5%, obtained in Moufida. Overall, the response in PC to the nitrogen application follows a linear regression pattern as shown in Figure 4, according to the equation: PC = (0.059 × nitrogen rate) + 36.868, with R2 = 0.936
 |
Fig. 3 Protein content variation in six rapeseed varieties under different nitrogen rates. |
 |
Fig. 4 Protein content in function of nitrogen application. |
3.4 Dry matter content
Analysis of variance revealed significant differences among nitrogen rates for dry matter content (DM) in rapeseed meal. However, the varieties studied were comparable and there was no significant effect of the interaction nitrogen rate by variety on this parameter (Tab. 3). The effect of nitrogen rate was positive on DM as this latter increased from 87.2% to 89.5% by increasing nitrogen application level from 0 to 120 kg N ha−1.
3.5 Neutral detergent fiber (NDF)
ANOVA analysis showed that NDF content in rapeseed meal was affected significantly only by genotype. For all nitrogen treatments combined, the smallest NDF content was observed in Lila exhibiting 26.2%, followed by Adila with 26.4%, while the highest NDF content was 28.4%, recorded in Trapper. However, the effect of nitrogen rate and nitrogen × variety interaction was not significant on NDF content (Tab. 3). In fact, a slight and non-significant decrease of about 1% was found in this parameter between N0 and N150 treatments.
3.6 Acid detergent fiber (ADF)
Highly significant differences among the nitrogen rates applied and significant differences among the varieties experimented were observed for ADF content. However, there was no significant effect of the interaction nitrogen rate × variety on this trait (Tab. 3). As shown in Table 3, ADF content of all varieties investigated decreased by increasing nitrogen application rate. In fact, ADF content decreased from 19%, without nitrogen fertilization, to 14.7%, recorded at 150 kg N ha−1 treatment. Regardless of the nitrogen rate, the varieties Lila and Alia had the lowest average ADF content (15.7%), while Narjisse and Moufida exhibited the highest one, with an average of 17.7%.
3.7 Acid detergent lignin (ADL)
Acid detergent lignin (ADL) content was significantly affected by nitrogen application level and genotypic variation. However, ADL content was not significantly influenced by the interaction nitrogen rate × genotype (Tab. 3). In all the varieties studied, ADL content decreased linearly from 7.5% for N0 to 5.4% for N150 treatment. With regard the varietal variation, the highest ADL content was observed in Moufida, with an average of 7.6%, while the lowest one was recorded in Narjisse, with a mean value of 6%.
3.8 Ash content
As shown in Table 3, significant differences among the nitrogen rates and among the varieties were noticed for RSM ash content (AC). Nevertheless, the effect of their interaction was not significant. Ash content in all varieties increased with the nitrogen application rate. The varieties Narjisse, Alia and Trapper reached their maximum AC at N120, while the other varieties registered their highest values at N150 (Fig. 5). The overall highest AC values were 11% and 11.1%, recorded at N150 and N120, respectively, whilst the lowest value was 7.5%, obtained for N0 (Tab. 3). Regardless of nitrogen rate applied, the variety Alia had the highest AC, with an average value of 10.8%, which is comparable to the varieties Lila and Adila having a mean content of 10.7% and 10.3%, respectively. However, the lowest AC was 7.7%, registered in Trapper.
 |
Fig. 5 Variation of ash content in six rapeseed varieties under different nitrogen rates. |
3.9 Correlations among the traits investigated
The coefficients of correlation among the different traits are shown in Table 4. Meal yield has a positive and significant correlation with dry matter (r = 0.46) and PC (r = 0.50). Also, PC is significantly and positively associated with dry matter (r = 0.38), but negatively with ADL (r = –0.51) and ADF (r = –0.37). Regarding fiber content, NDF is associated positively with ADF and ADL (r = 0.46 and r = 0.31, respectively). Nevertheless, the strongest correlation is found between ADF and ADL (r = 0.67). Finally, ash content is significantly and negatively correlated with ADF (r = –0.46), ADL (r = –0.31), and NDF (r = –0.30) (Tab. 4).
Correlation coefficient matrix for the different traits studied.
4 Discussion
We found that meal yield increased significantly by increasing the nitrogen rate. This was mainly due to a substantial improvement of seed yield as a result of nitrogen fertilization. Besides, it is well known that increasing nitrogen fertilization causes a decrease in seed oil content in rapeseed (Öztürk, 2010; Paye et al., 2021; Yahbi et al., 2022), which led to a concomitant increase in by-product (meal) content. Moreover, the absence of yield improvement and then of meal yield beyond N120 might be a result of the high temperature and the very low precipitation on the end of April and May which lead to maturity advancement. Also, genotype main effect significantly influenced meal yield. The hybrid variety Trapper exhibited the highest meal yield, compared to the rest of varieties. This can be explained by its highest performance in terms of seed yield. In fact, through our investigation of the effect of nitrogen fertilization on agronomic parameters, we did find that Trapper was the most productive for all the nitrogen rates applied (data not yet published).
PC is the most important trait that makes rapeseed meal one of the most interesting meals in the word. PC in meal was affected by nitrogen rate and variety. Our findings are in agreement with those of other authors who reported the positive response of meal PC to nitrogen treatment canola (Brennan and Bolland, 2007), and in rapeseed (Zuo et al., 2016). From 0 to N120, we recorded a substantial gain in mean PC of 8.34% which is markedly higher than the increase in PC of 2.1%, from 0 to 200 kg N ha−1, reported by Aminpanah (2013). This difference might be due to the genotypes experimented in both the studies and the contrasted environmental conditions under which those genotypes were planted and fertilized. Genotypic variation for crude PC in rapeseed meal was reported in previous studies (Olukosi et al., 2017; Watts et al., 2021). A variation between 39.49% in Moufida and 44.10% in Adila was observed in the present study, whereas in previous studies, it was reported that PC ranged from 36.7% to 43.9% (Olukosi et al., 2017) and from 35% to 40% (Cheng et al., 2022). As it was expected, nitrogen fertilization has a positive effect on PC that reached its highest value (44.9%) at N120 treatment. This increase in PC might be explained by the structural role of nitrogen in protein composition and the positive effect of nitrogen on photosynthates production and their translocation to seeds (Mehmet, 2008). no significant effect of the interaction nitrogen rate by variety on PC, indicating that the investigated varieties responded similarly or according to the same pattern to the nitrogen rates applied. In other words, the highest-performing varieties in terms of PC for low nitrogen application maintain their superiority also for high nitrogen application. In another side, protein yield per hectare as the PC multiplying by the MY, has been improved by the nitrogen rate. Increase the nitrogen rate from N0 to N120 increased the protein yield from 370.3 to 801.9 kg ha−1. With regard to rapeseed varieties it was Trapper who had the highest protein yield with 627.6 kg ha−1 followed by Narjisse and Alia with 624.9 and 615.2 kg ha−1 respectively. While the productivity of nitrogen application which is the amount of proteins gives a kg of nitrogen applied (kg protein kg N applied−1), it has decreased by increasing nitrogen rate (Fig. 6). This decrease can be explained by the fact that rapeseed plants are unable to absorb nitrogen at a high N rate because the supply of N is exceeding plant requirement and the possibility of N loss by leaching and denitrification.
Meal dry matter content was affected by nitrogen fertilization, with a significant increase from 87.2% for N0 to 89.5% for N120. This indicates that the higher the level of nitrogen fertilization, the lower the moisture content in rapeseed meal. On the other hand, the investigated varieties were found to be comparable for this parameter, with a narrow and non-significant variation from 87.9% in Trapper to 88.4% in Alia. These values are lower than those of Watts et al. (2021) who reported a dry matter content ranging from 88.6% to 91.1% across the ten rapeseed cultivars.
Neutral detergent fiber (NDF) content was found to be only affected by the genotype main effect. This imply that nitrogen fertilization has no impact on the content of NDF in rapeseed. Our result support the result observed by Costa et al. (2013), which showed that the NDF of rapeseed was similar under three nitrogen rates (0, 50 and 150 kg N ha−1). Moreover, a previous study on sorghum and black oat forages did also show that NDF content was similar under different nitrogen application rates (Restelatto, 2013), which is in agreement with our findings. NDF content can be affected not only by nitrogen level but also by other factor especially the method and the temperature of extraction (Pastuszewska et al., 2003; Kasprzak et al., 2016). On the contrary, significant differences were observed among the six varieties for NDF ranging from 26.2% in Lila to 28.4% in Trapper. This corroborates the findings of Dimov et al. (2012) having also reported that their rapeseed cultivars showed large differences for NDF content. Currently, NDF is an indicator for meal quality assessment in the feed industry, and an NDF content lower than 280 g kg−1 is considered for superior rapeseed meal quality (Wang et al., 2019). Apart from Trapper and Narjisse that showed a NDF content of 282.5 and 283.8 g kg−1, respectively, all the other varieties studied has a NDF content lower than 280 g kg−1, which makes them suitable for the production of high-quality rapeseed meal. Concerning acid detergent fiber (ADF) and acid detergent lignin (ADL), we found that their contents were significantly impacted by the nitrogen rate and the variety, but not by their interaction. The content of ADF dropped from 19% at N0 to 14.7% at N150 and that of ADL from 7.5% at N0 to 5.4% at N150. In concordance with our findings Costa et al. (2013) have found that the ADF and ADL decreased significantly with the increase of nitrogen supply from N0 to N150. Moreover, Sher et al. (2016) reported that as mentioned, and discussed before, nitrogen fertilization leads to an improvement of PC in rapeseed meal, which might cause a dilution effect for fibers content during seed filling. This can be confirmed by the negative correlation between PC and ADF (r = –0.37) and ADL (r = –0.51). Furthermore, this study showed that NDF remain constant while ADF varies, that means that hemicellulose in the seeds has changed. Genotypic variation for ADF and ADL was observed in this study. In fact, ADF content ranged from 15.7% in Lila to 17.7% in Narjisse, while ADL content varied from 6% in Narjisse to 7.6% in Moufida. These values are much lower than those of Dimov et al. (2012) who found a genotypic variation from 20.5% to 26.8% for ADF and from 8.5% to 13.2% for ADL. These differences can be explained by the genetic potential of the varieties used and by the effect of the contrasting environments under which those varieties were growing. Moreover, For the fiber content it has showed that the nitrogen fertilization had a significant effect on ADF and ADL but not on NDF. In a previous study Carré et al. (2016) reported that in rapeseed the high fiber content of whole seeds is clearly driven by the presence of hulls, which held the main content of lignin (Hulls contained 73% of the NDF; 80% of the ADF, 95% of the lignin). So it seems that nitrogen fertilization probably has more influence on the filling of the embryo than on the formation of the integument. Furthermore, the ADF and NDF in the “Double-low” rapeseed meal is affected more by the varieties, whose content vary among different varieties, and the coefficients of variance is reportedly by 29.3% and 30.8%, respectively (Peng and Jiang, 1999). Moreover, the NDF fraction consists predominantly of hemicellulose, cellulose and lignin, whereas the ADF fraction comprises cellulose and lignin. ADL represents the non-digestible lignin fraction. Lignin is the predominant component of the seed coat, followed by cellulose and hemicellulose (Carré et al., 2016).
Finally, our study revealed that nitrogen rate and variety have also affected the ash content in rapeseed meal. An increase of ash content was observed following the elevating nitrogen application rates, from 7.5% in the absence of nitrogen fertilization to 11.1% at N120 fertilization. This increase could be a result of the effect of nitrogen on seed composition. In fact, rapeseed nitrogen fertilization help increase the content of micronutrients, such as P, K, Zn, Cu and Fe, in seeds (Fordoński et al., 2016). Our result showed that the ash content found is bigger than the values shown in Table 1 (6.8% and 7% in rapeseed and canola meal respectively), these differences might be due to the varieties characteristics and meal composition. Moreover, the German cultivar Trapper has ashes content (7.7%) which is close to the ash content mentioned in Table 1 (European level) while the other ones are Moroccan. It is possible that they share a common parent with great ability to store minerals.
In addition to meal yield and PC associated with high production of proteins per hectare, is the most important and desirable meal quality index. Accordingly, the variety Trapper seems to be the most interesting for rapeseed meal-based animal feed. To optimize the quantity and quality of its meal production, this variety should be grown under 120 kg N ha−1 conditions. Due to the inverse relation between PC and overall fiber content, mainly ADL and ADF, explained by the negative correlation, breeding for high PC would enable improving the rapeseed meal nutritional value by reducing the fiber content. Negative association between PC and overall fiber content, including NDF, ADF and ADL, was also reported in a previous study (Dimov et al. (2012)).
 |
Fig. 6 Effect of nitrogen rate on productivity of nitrogen application rate. |
5 Conclusion
All the traits investigated were found to be affected by nitrogen fertilization, except for the NDF content. As expected, a positive effect of nitrogen application rate on meal yield was revealed. Also, dry matter, ash content and PC were increased by raising the nitrogen rate to reach their peak at 120 kg N ha−1. On the contrary, this study has shown for the first time that ADF and ADL content in rapeseed meal decreased as a result of increasing nitrogen fertilization dose. Therefore, nitrogen fertilization could improve the nutritional value of rapeseed meal, using a quite high application rate, namely 120 kg N ha−1 for our conditions. Moreover, a significant variation among the varieties studied was found for all the parameters investigated. The variety Trapper recorded the highest MY, PC and protein yield, with a protein yield over of 727 kg ha−1. So, Trapper cultivated under N120 fertilization conditions could be recommended to produce high-quality rapeseed meal to feed livestock. Additional studies in other environments (different sites and years) are needed to investigate their main effect on chemical composition of rapeseed meal and to assess their interaction with the varieties and nitrogen fertilization rates.
Conflicts of Interest
The authors have declared that no competing interests exist.
Author contribution statement
Conceptualization: Mohammed Yahbi, Nabila El Alami, Abdelghani Nabloussi, Khalid Daoui. Data curation: Not applicable. Formal analysis: Mohammed Yahbi, Nabila El Alami, Abdelghani Nabloussi, Khalid Daoui. Investigation: Mohammed Yahbi. Methodology: Abdelhafid Keli. Project administration: Nabila El Alami, Abdelghani Nabloussi, Khalid Daoui. Supervision: Nabila El Alami, Abdelghani Nabloussi, Khalid Daoui. Visualization: Nabila El Alami, Abdelghani Nabloussi, Khalid Daoui, Abdelhafid Keli. Writing − original draft: Mohammed Yahbi. Writing − review & editing: Nabila El Alami, Abdelghani Nabloussi, Khalid Daoui, Abdelhafid Keli, Abdelwahed Maataoui.
References
- Adewole DI, Rogiewicz A, Dyck B, Slominski BA. 2016. Chemical and nutritive characteristics of canola meal from Canadian processing facilities. Anim Feed Sci Technol 222: 17–30. [CrossRef] [Google Scholar]
- Ahmad G, Jan A, Arif M, Jan, MT, Khattak, RA. 2009. Influence of nitrogen and sulfur fertilization on quality of canola (Brassica napus L.) under rainfed conditions. Journal of Zhejiang University Science, 8, 731-737. [Google Scholar]
- Aksouh-Harradj NM, Campbell LC, Mailer RJ. 2006. Canola response to high and moderately high temperature stresses during seed maturation. Canadian journal of plant science, 86(4), 967-980. https://doi.org/10.4141/P05-130. [Google Scholar]
- Aminpanah H. 2013. Effect of nitrogen rate on seed yield, protein and oil content of two canola (Brassica napus L.) cultivars. Acta Agric Slov 101: 183–190. [CrossRef] [Google Scholar]
- Asare E, Scarisbrick DH. 1995. Rate of nitrogen and sulphur fertilizers on yield, yield components and seed quality of oilseed rape (Brassica napus L.). Field Crops Res 44: 41–46. [CrossRef] [Google Scholar]
- Badani AG, Snowdon RJ, Wittkop B, et al. 2005. Colocalization of a partially dominant gene for yellow seed colour with a major QTL influencing acid detergent fibre (ADF) content in different crosses of oilseed rape (Brassica napus). Genome 49: 1499–1509. [Google Scholar]
- Bell JM. Meal and by-product utilization in animal nutrition. In Kimber D, McGregor DI, eds. Brassica oilseeds, production and utilization. Oxon, UK: CAB International, 1995, pp. 301–337. [Google Scholar]
- Bellostas N, Sorensen H, Sorensen S. Quality of rapeseed meal for animal nutrition and as a source of value-added products-glucosinolates, protein and fibers. In: letin Nutrition and Processing Workshop of the 12th International Rapeseed Congress held in Wuhan, China ( 26–30 March 2007). 2007, p. 24. [Google Scholar]
- Bjergegaard C, Eggum BO, Jensen SK, Sørensen H. 1991. Dietary fibres in oilseed rape: physiological and antinutritional effects in rats of IDF and SDF added to a standard diet. J Anim Physiol Anim Nut 66: 6979. [Google Scholar]
- Bojanowska M. 2017. Changes in chemical composition of rapeseed meal during storage, influencing nutritional value of its protein and lipid fractions. J Anim Feed Sci 26: 157–164. [CrossRef] [Google Scholar]
- Carré P, Citeau M, Robin G, Estorges M. 2016. Hull content and chemical composition of whole seeds, hulls and germs in cultivars of rapeseed (Brassica napus). OCL 23: A302. [CrossRef] [EDP Sciences] [Google Scholar]
- Costa D, Santos CM, Silva A, Rodrigues, MA. 2013. The response of rapeseed cultivar hydromel to nitrogen fertilization: chemical composition and antioxidant capacity. Poster in XXIII Encontro Nacional da SPQ, que decorreu de 12 a 14 de junho de 2013) na universidade de Aveiro. [Google Scholar]
- Dimov Z, Suprianto E, Hermann F, Möllers C. 2012. Genetic variation for seed hull and fibre content in a collection of European winter oilseed rape material (Brassica napus L.) and development of NIRS calibrations. Plant Breed 131: 361–368. [CrossRef] [Google Scholar]
- Brennan RF, Bolland MDA. 2007. Effect of fertiliser phosphorus and nitrogen on the concentrations of oil and protein in grain and the grain yield of canola (Brassica napus L.) grown in south-western Australia. Aust J Exp Agric 47: 984–991. [CrossRef] [Google Scholar]
- Cheng H, Liu X, Xiao Q, et al. 2022. Rapeseed meal and its application in pig diet: a review. Agriculture 12: 849. [CrossRef] [Google Scholar]
- Enami HR. 2011. A review of using canola/rapeseed meal in aquaculture feeding. J Fish Aquat Sci 6: 22–36. [Google Scholar]
- FEEDTABLES. 2023.xxx Available from: https://feedtables.com/content/table-feed-profile?feed_pr_id=222075 (last consult: 2023/02/11). [Google Scholar]
- Fordoński G, Pszczółkowska A, Okorski A, Olszewski J, Załuski D, Gorzkowska A. 2016. The yield and chemical composition of winter oilseed rape seeds depending on different nitrogen fertilization rates and preceding crop. Elem J 21: 1225–1234. [Google Scholar]
- Hammac WA, Maaz TM, Koenig RT, Burke IC, Pan WL. 2017. Water and temperature stresses impact canola (Brassica napus L.) fatty acid, protein, and yield over nitrogen and sulfur. J Agric Food Chem 65: 10429–10438. [CrossRef] [PubMed] [Google Scholar]
- Hao X, C. Chang, GJ, Travis. 2004. Short communication; Effect of long-term cattle manure application on relations between nitrogen and oil content in canola seed. Journal of Plant Nutrition and Soil Science, 167: 214–215. [Google Scholar]
- Hannoufa A, Pillai BV, Chellamma S. 2014 Genetic enhancement of Brassica napus seed quality. Transgenic Res 23: 39–52. [CrossRef] [PubMed] [Google Scholar]
- Hasan M, Friedt W, Pons-Kühnemann J, Freitag NM, Link K, Snowdon RJ. 2008. Association of gene-linked SSR markers to seed glucosinolate content in oilseed rape (Brassica napus ssp. napus). Theor Appl Genet 116: 1035–1049. [CrossRef] [PubMed] [Google Scholar]
- Hossain Z, Johnson EN, Wang L, Blackshaw RE, Gan Y. 2019. Comparative analysis of oil and protein content and seed yield of five Brassicaceae oilseeds on the Canadian prairie. Ind Crops Prod 136: 77–86. [CrossRef] [Google Scholar]
- Huang XQ, Huang T, Hou GZ, Li L, Hou Y, Lu YH. 2016. Identification of QTLs for seed quality traits in rapeseed (Brassica napus L.) using recombinant inbred lines (RILs). Euphytica 210: 1–16. [Google Scholar]
- Hüsken A, Baumert A, Strack D, Becker HC, Möllers C, Milkowski C. 2005. Reduction of sinapate ester content in transgenic oilseed rape (Brassica napus) by dsRNAi-based suppression of BnSGT1 gene expression. Mol Breed 16: 127–138. [CrossRef] [Google Scholar]
- Kalogianni AI. Moschovas M, Chrysanthakopoulou F et al. 2022. Theeffects of replacing soybean meal with rapeseed meal, cottonseed cake, and fava beans on the milk yield and quality traits in milking ewes. Animals 12: 274. [Google Scholar]
- Kenny KY So, Robert WD. 2021. Breeding Canola (Brassica napus L.) for Protein in Feed and Food. Plants, 10(10). https://doi.org/10.3390/plants10102220 [Google Scholar]
- Kasperzak MM, Houdijk JGM, Kightley S, et al. 2016. Effects of rapeseed variety and oil extraction method on the content and ileal digestibility of crude protein and amino acids in rapeseed cake and softly processed rapeseed meal fed to broiler chickens. Anim Feed Sci Technol 213: 90–98. [CrossRef] [Google Scholar]
- Khajali F, Slominski BA. 2012. Factors that affect the nutritive value of canola meal for poultry. Poult Sci 91: 2564–2575. [CrossRef] [PubMed] [Google Scholar]
- Lin Y, Watts DB, Torbert HA, Howe JA. 2020. Influence of nitrogen rate on winter canola production in the southeastern influence of nitrogen rate on winter canola production in the southeastern United States. Agron J 112: 2978–2987. [CrossRef] [Google Scholar]
- Malabat C, Atterby H, Chaudhry Q, Renard M, Guéguen J. Genetic variability of rapeseed protein composition, in 11th International Rapeseed Congress − Enhanced Value of Cruciferous Oilseed Crops by Optimal Production and Use of the High Quality Seed Components, Copenhagen, 2003, pp. 205–208. [Google Scholar]
- Matthäus B. 2002. Antioxidant activity of extracts isolated from residues of oilseeds, such as rapeseed or sunflower. J Agric Food Chem 13: 22–25. [Google Scholar]
- Mustafa AF, Christensen DA, Mckinnon JJ, Newkirk R. 2000. Effects of stage of processing of canola seed on chemical composition and in vitro protein degradability of canola meal and intermediate products. Can J Anim Sci 80: 211–214. [CrossRef] [Google Scholar]
- Narits L. 2010. Effect of nitrogen rate and application time to yield and quality of winter oilseed rape (Brassica napus L. var. oleifera subvar. biennis). Agron Res 8: 671–686. [Google Scholar]
- Nesi N, Delourme R, Brégeon M, Falentin C, Renard M. 2008. Genetic and molecular approaches to improve nutritional value of Brassica napus L. seed. C R Biol 331: 763–771. [CrossRef] [PubMed] [Google Scholar]
- Ochodzki P, Rakowska M, Bjergegaard C, Sorensen H. 1995. Studies on enzymatic fractional, chemical composition and biological effects of dietary fibre in rape seed (Brassica napus L.) 1. Chemical composition of seeds and characteristics of soluble and insoluble dietary fibre of spring and winter type variety. J Anim Feed Sci 4: 127–138. [CrossRef] [Google Scholar]
- Olukosi OA, Kasprzak MM, Kightley S, Carre P, Wiseman J, Houdijk JGM. 2017. Investigations of the nutritive value of meals of double-low rapeseed and its influence on growth performance of broiler chickens. Poult Sci 96: 3338–3350. [CrossRef] [PubMed] [Google Scholar]
- Öztürk Ö. 2010. Effects of source and rate of nitrogen fertilizer on yield, yield components and quality of winter rapeseed (Brassica napus L.). Chil J Agric Res 70: 132–141. [Google Scholar]
- Paye WS, Singh P, Begna S, et al. 2021. Winter canola yield and nitrogen use efficiency in a semiarid irrigated condition. Agron 113: 2053–2067. [CrossRef] [Google Scholar]
- Peng J, Jiang S. 1999. Effects of dietary fibre on digestibility and availability of amino acids in pig. Pigs Poultry 5: 10–12 ( in Chinese). [Google Scholar]
- Peng J. 2000. Evaluation and improvement of quality of Chinese double-low rapeseed meal. The Ph.D dissertation of Huazhong Agricultural University (in Chinese). xx [Google Scholar]
- Peyronnet C, Alibert L, Bouvarel I, Bird I. 2010. Pois et tourteau de colza: deux matières premières complémentaires pour l’alimentation animale. OCL J 17: 319–324. [CrossRef] [EDP Sciences] [Google Scholar]
- Raab B, Leman H, Schwenke KD, Kozlowska H. 1992. Comparative study of the protein patterns of some rapeseed (Brassica napus L.) varieties by means of polyacrylamide gel electrophoresis and high-performance liquid chromatography. Food/Nahrung 36: 239–247. [CrossRef] [Google Scholar]
- Rathke GW, Christen O, Diepenbrock W. 2005. Effects of nitrogen source and rate on productivity and quality of winter oilseed rape (Brassica napus L.) grown in different crop rotations. Field Crops Res 94: 103–113. [CrossRef] [Google Scholar]
- Restelatto R, Pavinato PS, Sartor LR, Paixão SJ. 2013. Production and nutritional value of sorghum and black oat forages under nitrogen fertilization. Grass Forage Sci 69: 693–704. [Google Scholar]
- Ribaillier D, Delorme, S, Bureau J. 1995. Changes in agronomic pratices, in relation with the new european agricultural politic, can leads to a decrease of protein level in the rapeseed seeds, related to. Proteins (N x 6, 25), 4(6), 34-9. Proceedings of 9th International Rapeseed Congress, Cambridge. 3: 870–872. [Google Scholar]
- Sanjayan N, Heo JM, Nyachoti CM. 2014. Nutrient digestibility and growth performance of pigs fed diets with different levels of canola meal from Brassica napus black and Brassica juncea yellow. J Anim Sci 92: 3895–3905. [CrossRef] [PubMed] [Google Scholar]
- Schatzki J, Ecke W, Becker HC, Möllers C. 2014. Mapping of QTL for the seed storage proteins cruciferin and napin in a winter oilseed rape doubled haploid population and their inheritance in relation to other seed traits. Theor Appl Genet 127: 1213–1222. [CrossRef] [PubMed] [Google Scholar]
- Sher A, Hassan FU, Ali H, Hussain M, Sattar A. 2016. Enhancing forage quality through appropriate nitrogen dose, seed rate and harvest stage, in sorghum cultivars grown in Pakistan. Grassland Science, 63(1), 15-22. https://doi.org/10.1111/grs.12137 [Google Scholar]
- Slominski BA, Jia W, Rogiewicz A, Nyachoti CM, Hickling D. 2012. Low-fiber canola. Part 1. Chemical and nutritive composition of the meal. J Agric Food Chem 60: 12225–12230. [CrossRef] [PubMed] [Google Scholar]
- Theodoridou K, Yu P. 2012. Effect of processing conditions on the nutritive value of canola meal and presscake. Comparison of the yellow and brown-seeded canola meal with the brown-seeded canola presscake. J Sci Food Agric 93: 1986–1995. [Google Scholar]
- Ton LB, Neik TX, Batley J. 2020. The use of genetic and gene technologies in shaping modern rapeseed cultivars (Brassica napus L.). Genes 11: 1161. [CrossRef] [PubMed] [Google Scholar]
- Van Soest, PJ, Robertson JB, Lewis BA. 1991. Methods of dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J Dairy Sci 74: 3583–3597. [CrossRef] [PubMed] [Google Scholar]
- Wang Y, Liu J, Wei F, Liu X, Yi C, Zhang Y. 2019. Improvement of the nutritional value, sensory properties and bioavailability of rapeseed meal fermented with mixed microorganisms. Lwt 112: 108238. [CrossRef] [Google Scholar]
- Watts ES, Rose SP, Mackenzie AM, et al. 2021. Investigations into the chemical composition and nutritional value of single-cultivar rapeseed meals for broiler chickens. Arch Anim Nutr 75: 209–221. [CrossRef] [Google Scholar]
- Weier D, Mittasch J, Strack D, Milkowski C. 2007. The genes BnSCT1 and BnSCT2 from Brassica napus encoding the final enzyme of sinapine biosynthesis: molecular characterization and suppression. Planta 227: 375–385. [CrossRef] [PubMed] [Google Scholar]
- Wittkop B, Snowdon RJ, Friedt W. 2009. Status and perspectives of breeding for enhanced yield and quality of oilseed crops for Europe. Euphytica 170: 131–140. [Google Scholar]
- Yahbi M, Nabloussi A, Maataoui A, El Alami N, Boutagayout A, Daoui K. 2022. Effects of nitrogen rates on yield, yield components, and other related attributes of different rapeseed (Brassica napus L.) varieties. OCL J 29: 8. [CrossRef] [EDP Sciences] [Google Scholar]
- Zhao J, Becker HC, Zhang D, Zhang Y, Ecke W. 2005. Oil content in a European× Chinese rapeseed population: QTL with additive and epistatic effects and their genotype-environment interactions. Crop Sci 45: 51–59. [CrossRef] [Google Scholar]
- Zhao J, Becker HC, Zhang, D, ZhangY, Ecke W. 2006. Conditional QTL mapping of oil content in rape- seed with respect to protein content and traits related to plant development and grain yield. Theoretical and Applied Genetics, 113,33–38. https://doi.org/10.1007/s00122-006-0267-5 [Google Scholar]
- Zuo QS, Zhou GS, Yang SF, et al. 2016. Effects of nitrogen rate and genotype on seed protein and amino acid content in canola. J Agric Sci 154: 438–455. [CrossRef] [Google Scholar]
Cite this article as: Yahbi M, Keli A, El Alami, N, Nabloussi A, Maataoui A, Daoui K. 2024. Chemical composition and quality of rapeseed meal as affected by genotype and nitrogen fertilization. OCL 31: 5.
All Tables
Main effect of nitrogen rate and variety and their interaction on meal yield and its chemical proprieties of RSM as fed (%). (Data under the nitrogen fertilizer treatment were averaged across varieties, and data under the variety treatment are averaged across fertilizer levels.)
All Figures
|
Fig. 1 Monthly precipitations (bars) and air temperatures (lines) recorded in the experimental station during the cropping season. |
| In the text | |
|
Fig. 2 Meal yield in six rapeseed varieties as affected by nitrogen rate. |
| In the text | |
|
Fig. 3 Protein content variation in six rapeseed varieties under different nitrogen rates. |
| In the text | |
|
Fig. 4 Protein content in function of nitrogen application. |
| In the text | |
|
Fig. 5 Variation of ash content in six rapeseed varieties under different nitrogen rates. |
| In the text | |
|
Fig. 6 Effect of nitrogen rate on productivity of nitrogen application rate. |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.